|
|
| INTEGRATORI |
| NUTRIZIONE |
| BIOLOGICO |
|
|
| COMPOSIZIONE ALIMENTI |
| ALLENAMENTO |
| Oltre 80 consigli su alimentazione e allenamento con i pesi |
| PRODOTTI TIPICI D'ITALIA |
| RICETTE |
| Tante ricette dietetiche per Voi! |
| Evoluzione degli Antiossidanti nella alimentazione: Ruolo dello Iodio |
|
|||
|
|
di Sebastiano Venturi, Mattia Venturi . L’atomo di iodio (Simbolo I; P.A.126.9; N.A 53) è costituito da 53 elettroni, 53 protoni e 74 neutroni, e rappresenta una sorta di “cenere nucleare”. Infatti, deriva da un processo di nucleo-sintesi avvenuto più di 10 miliardi di anni fa’ in una stella-supernova, che esplodendo lo ha disperso nel pulviscolo primordiale, il quale condensandosi, circa 5 miliardi di anni fa’, ha formato il nostro pianeta-Terra. Lo iodio è uno degli atomi più ricchi di elettroni presenti nel nostro corpo ed è indispensabile nella dieta di tutti gli esseri viventi animali. Lo iodio (I) è scarsamente reperibile nella superficie terrestre, perché nel corso di centinaia di milioni di anni è stato dilavato, da piogge e glaciazioni, e trasportato dalla crosta terrestre verso il mare, il quale si è arricchito progressivamente di iodio, sotto forma di ioduri (I-) e di iodati. |
|||
|
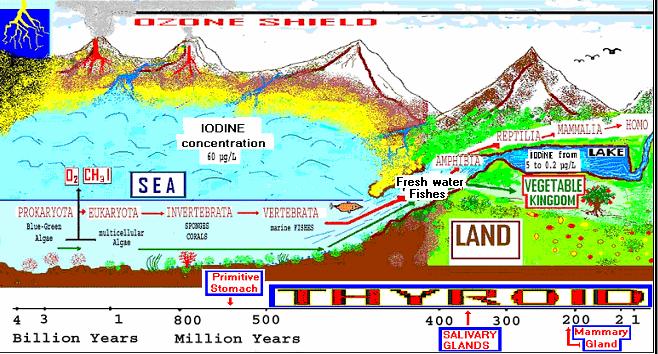
Fig 1. Struttura atomica dello iodio Infatti le acque marine sono ricche di iodio, circa 50-60 microgrammi (mg) per litro, mentre le acque terrestri: estuari, fiumi, laghi, ne contengono quantità da 10 a 200 volte inferiori ( 5 - 0.2 mg/L). Una piccola parte di iodio evapora nell’aria (anche come iodo-metano gassoso) e precipita nel suolo con le piogge, soprattutto in vicinanza delle zone costiere. Il ciclo geo-biologico dello iodio, è in parte simile a quello del selenio. Lo iodio viene captato dalle cellule come ioduro (I-) soprattutto tramite il NIS (sodium iodide symporter), ma anche altri trasportatori sono oggi stati identificati. Il NIS è il trasportatore glicoproteico transmembrana dello ioduro, la cui molecola, nell’uomo, è stata clonata e caratterizzata da Dai e coll.(1996) e Smanik e coll.(1996). Essendo presumibilmente molto antico, il NIS è poco specifico e secondo Wolff (1964) non è in grado di distinguere lo ioduro da altri atomi o piccole molecole, come i nitrati, i nitriti, i fluoruri, i tiocianati, i pertecnati ecc., aventi stessa carica elettrica e simili dimensioni atomiche o molecolari, che sono “in competizione” con lo ioduro, comportandosi come “pseudo-ioduri”. La caratteristica elettrochimica dello iodio è quella di attirare e cedere facilmente un elettrone (con un potenziale redox di -0.54 Volt). Questa proprietà lo rende un efficiente donatore-accettore di elettroni, che è una delle caratteristiche fondamentali delle sostanze antiossidanti. Infatti:
2 I- à I2 + 2 e- (elettroni) = - 0.54 Volt ; 2 I- + Perossidasi + H2O2 + 2 Tirosina à 2 Iodio-Tirosina + H2O + 2 e- (antiossidanti) ; 2 e- + H2O2 + 2 H+ (della soluzione acquosa intracellulare) à 2 H2O Tab. A 2 I- + Peroxidase + H2O2 + Tyrosine, Histidine, Lipids, Carbons à à Iodo-Compounds + H2O + 2 e- (antioxidants)
Iodo-Compounds: Iodo-Tyrosine, Iodo-Histidine, Iodo-Lipids, Iodo-Carbons
Tab. B Tab. A e Tab. B. Meccanismi biochimici antiossidanti degli ioduri, probabilmente uno dei più antichi meccanismi di difesa dai radicali liberi dell’ossigeno, già presenti nei Cianobatteri circa 3 miliardi e mezzo di anni fa’ (Venturi, 1985)
Le alghe marine sono in grado, tramite enzimi alo-perossidasici, di catalizzare l’incorporazione dello ioduro in alcuni idrocarburi producendo iodio-metano gassoso (CH3I) e altri alo-idrocarburi, nella atmosfera. Secondo Petersén (1996), Colin et al (2003), Gall et al., (2003) Kuepper e coll. (1998) questa produzione è il risultato della primitiva fotosintesi, della produzione di ossigeno e della respirazione cellulare, iniziate oltre tre miliardi di anni fa’; ed è dovuta allo scopo di ridurre il danno dei radicali liberi dell’ossigeno (ROS), come il perossido di idrogeno (H2O2), i superossidi ed i radicali ossidrilici. Recentemente una altra via metabolica è stata descritta, tramite la quale lo ioduro viene incorporato negli acidi grassi poli-insaturi (PUFA) delle membrane cellulari, proteggendoli dalle perossidazioni (Cocchi, Venturi, 2000). Sia le cellule tiroidee che quelle di altri tessuti I-captanti, come le cellule della mucosa gastrica e delle ghiandole salivari ecc., sono in grado di produrre "in vitro" mono-iodio-tirosina (MIT) e di-iodio-tirosina (DIT) legate a proteine e anche alcuni poco conosciuti iodio-lipidi, che sembrano avere una importante funzione strutturale e metabolica come secondi messaggeri. In particolare il delta-iodiolattone (acido 6-iodio-5-idrossi-eicosatrienoico) è un potente inibitore della proliferazione delle cellule tiroidee e secondo Cann e coll. ( 2000) e Venturi (2001) gioca anche un ruolo nel controllo antiproliferativo dei tessuti extratiroidei I-concentranti.
Fig. 2. Iodio ed evoluzione. Più di tre miliardi di anni fa’ le alghe verdi-azzurre furono le prime cellule procariote a produrre ossigeno (allora tossico) e iodio-composti, tra cui iodo-metano (CH3I) gassoso, nella atmosfera terrestre. Da circa 800-700 milioni di anni la tiroxina (T4) è presente nell’esoscheletro degli invertebrati marini (spugne, coralli, conchiglie ecc.) senza possedere alcuna conosciuta azione ormonale. Circa 500-400 milioni di anni fa’, alcuni primitivi cordati iniziarono a risalire dal mare (ricco di iodio) le acque I-carenti degli estuari e poi dei fiumi. Circa 400-300 milioni di anni fa’ alcuni di questi primitivi vertebrati cominciarono ad evolversi in anfibi e poi in rettili, che poi popolarono permanentemente l’ habitat terrestre I-carente. Allora questi vertebrati terrestri ebbero bisogno di un nuovo efficiente organo dove poter accumulare il poco iodio presente nell’ habitat terrestre: il follicolo “tiroideo”. I vertebrati cominciarono poi ad utilizzare la T4 come trasportatore nelle cellule periferiche dello ioduro antiossidante, ed in seguito iniziarono a utilizzare la T3, grazie a i suoi nuovi recettori. La T3 divenne così l’ormone attivo nella metamorfosi e nella termogenesi, per un migliore adattamento al nuovo habitat terrestre (Venturi, 2004).
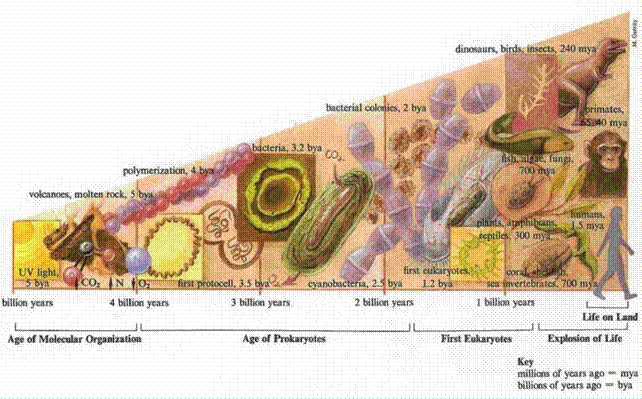
L’organismo umano contiene circa 25-50 milligrammi di iodio, di cui meno di 10-15 mg sono presenti nei follicoli della tiroide e meno di 1 mg negli ormoni tiroidei circolanti. La maggior parte, il 60-70 % di tutto lo iodio del corpo umano, è presente in sede extratiroidea ed è captato da diversi organi non-follicolari: stomaco, epidermide, mammella, ghiandole salivari, arterie, timo ecc. in cui, sembra ormai accertato, svolge una azione diretta antiossidante, non ormonale, ancora poco conosciuta. Tale azione, era già presente, secondo Venturi (1985), Petersen e coll. e Kuepper e coll. più di tre miliardi di anni fa nelle alghe verdi-azzurre (cianobatteri) e probabilmente ha costituito uno dei più antichi meccanismi antiossidanti di difesa dai ROS. Infatti queste alghe, ricche di iodio, furono le prime a produrre ossigeno, fino ad allora assente nella atmosfera terrestre. Per cui la cellula algale doveva possedere degli antiossidanti, efficaci e facilmente reperibili, per difendersi dalla tossicità dell’ossigeno. Gli ioduri, ed il selenio, diffusi e ben reperibili nelle acque marine, hanno avuto in ciò un ruolo determinante. Infatti, il selenio è presente nelle perossidasi e nelle deiodasi intracellulari, le quali sono capaci di estrarre elettroni dagli ioduri, e queste ultime gli ioduri dalle iodio-tironine. La vita nel nostro pianeta-Terra è iniziata nel mare circa 4 miliardi di anni fa’, e per tre miliardi e mezzo di anni è stata esclusivamente marina, solo negli ultimi 300-400 milioni di anni fa’, alcuni esseri viventi, protetti dai raggi ultravioletti solari dallo scudo dell’ozono (O3), iniziarono ad emergere dalle acque marine e ad abitare la terraferma (carente di iodio): prima i vegetali poi gli animali.
Fig. 3. La vita nel nostro pianeta è iniziata nel mare circa 4 miliardi di anni fa’ e per tre miliardi e mezzo di anni è stata esclusivamente marina, solo negli ultimi 300-400 milioni di anni, alcuni esseri viventi iniziarono ad emergere dalle acque marine e ad abitare la terraferma carente di iodio e di altri antiossidanti marini.
Si creò allora una grave crisi nutrizionale di iodio, ma anche di selenio e di altri meno conosciuti antiossidanti “marini”. Infatti, mentre nel mare tutti gli esseri viventi, potevano utilizzare lo iodio ed il selenio, con il trasferimento sulla terraferma si è interrotta la catena alimentare nutrizionale marina che li trasferiva (insieme agli acidi grassi omega-3), dal fitoplancton fino ai pesci marini. I vegetali “terrestri” hanno superato questa crisi nutrizionale di antiossidanti marini, perfezionando ed utilizzando sostanze antiossidanti alternative, alcune già elaborate negli estuari I-carenti, come i polifenoli, i flavonoidi, l’acido ascorbico, i carotenoidi, i tocoferoli ecc. di cui alcuni sono diventati fattori “vitaminici”, essenziali per l’uomo, come le vitamine C, A, E ecc. Infatti lo iodio non divenne più indispensabile per diverse specie vegetali terrestri. Alcuni antiossidanti ebbero una evoluzione “filogenetica” continua fino alle più recenti sostanze antiossidanti come il licopene e molti polifenoli, carotenoidi ecc. che si sono sviluppati solo recentemente nei pigmenti colorati antiossidanti presenti nei fiori e nella frutta delle piante angiosperme ( derivate dalle più antiche piante gimnosperme) che sono comparse sulla terra solo da circa 200-100 milioni di anni e sono divenute oggi il tipo di piante più numeroso. Gli animali, invece, hanno superato questa crisi nutrizionale, cercando anche di migliorare e di ottimizzare le scarse quantità di iodio disponibili sulla terraferma, mediante 3 meccanismi adattativi:
1) la creazione del follicolo tiroideo;
2) l’utilizzazione della Tiroxina (T4) come trasportatore di ioduri;
3) la formazione dei recettori della T3 e quindi della sua “nuova” funzione ormonale.
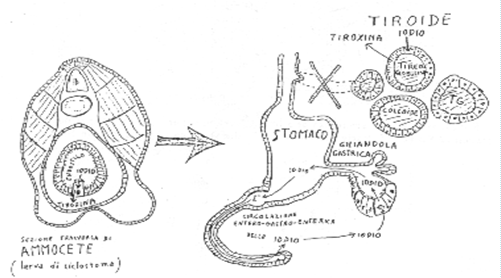
La somiglianza tra stomaco e tiroide è dovuta proprio alla comune filogenesi ed embriogenesi, essendo le cellule “tiroidee” derivate proprio dall’intestino primitivo, che era ed è capace di captare iodio e formare composti iodati. Questo spiega le comuni caratteristiche tra cui: la polarità e i microvilli apicali, la capacità di captare e di secernere iodio, la secrezione di ormoni aminoacidici e di simili glicoproteine (tireoglobulina e mucina) e inoltre la capacità di digerire tramite peptidasi e di riassorbire (tireoglobulina e cibo) ed infine i comuni antigeni di membrana e le malattie immunologiche associate.
Fig. 4. Metamorfosi dell’ammocete (larva di ciclostoma) in lampreda adulta, con neoformazione del follicolo "tiroideo" derivato dalle cellule gastroenteriche della larva. La formazione del follicolo-deposito di iodio sembra sia originata negli estuari, in preparazione della migrazione nelle acque dolci (iodo-carenti) dei fiumi terrestri. (da Magni M.A, 1985).
1) La formazione del follicolo tiroideo, che origina infatti dall’intestino I-captante primitivo, come efficiente forma di deposito dello iodio (iniziato nei primitivi cordati, sembra negli estuari, prima di migrare nelle acque interne terrestri I-carenti). E’ proprio grazie al deposito-riserva di iodio nei follicoli tiroidei, che noi uomini possiamo vivere per molte settimane senza assumere iodio e senza avere sintomi clinici di carenza.
2) La utilizzazione della T4, che non è , come gli ioduri, in competizione (a livello del NIS) con gli anioni monovalenti vegetali, ma ha un diverso, più moderno e specifico recettore. Infatti, la dieta vegetale terrestre è ricca di antagonisti dello ioduro sul NIS come i nitrati, nitriti, tiocianati, cianati, glicosidi ecc. sviluppatisi come strategia di difesa antiparassitaria. Questo nuovo meccanismo della T4 si è integrato, senza sostituirlo, a quello più antico del trasportatore dello ioduro (NIS), che è sempre funzionante, come si può ben vedere anche nelle I-scintigrafie total-body sotto riportate. La tiroxina, che prima veniva spesso secreta ed eliminata dalla cellula “marina”, diventa così un trasportatore endocellulare dello ioduro, molto più efficiente, come hanno dimostrato Evans e coll. Ricerche di Tseng e Latham e di Oziol e coll. hanno documentato, inoltre, un potere antiossidante ed inibitore della perossidazione lipidica della T4 e della rT3 (ma non della T3) superiore alle vitamine C ed E ed al glutatione; e Virgili e coll. hanno riportato che il trattamento con tiroxina protegge dai danni perossidativi intestinali indotti dalla carenza di zinco nei ratti. Inoltre gli ioduri difendono le cellule cerebrali dai danni perossidativi nei ratti ( Katamine, 1985) e sono stati utilizzati nella terapia di molte malattie umane degenerative su base perossidativa come arteriosclerosi, vasculopatie, artrosi ecc. in numerosi studi clinici degli anni ’50 in Europa, in cui a differenza degli USA, non era allora praticata la iodioprofilassi. Recenti studi stanno oggi evidenziando le basi biochimiche della azione antiossidante degli ioduri (Winkler e coll). Secondo Kahaly (2000) e Hak e coll.(2000) lo iodio e la funzionalità tiroidea sono importanti nel metabolismo dei lipidi, del colesterolo e nel ridurre l’aterosclerosi e l’ipertensione, mentre l’ipotiroidismo anche subclinico è oggi ritenuto causa importante di morbilità cardiovascolare. Recentemente Cann (2006) ha pubblicato una importante review su “iodio e malattie cardio-vascolari” riportandone numerose esperienze di efficacia preventiva e terapeutica. La I-concentrazione nella parete elastica della aorta presente ancora dopo 14 giorni dalla somministrazione di radio-ioduro-131 , visibile nella fig. 9 (in basso) ci fornisce anche, insieme alle proprietà antiossidanti dello ioduro, un razionale di questa azione anti-aterosclerotica. Venturi ha in corso di pubblicazione lavori su “ioduro come antiossidante e patologia orale” e sul ruolo dello iodio nel contesto generale della evoluzione degli antiossidanti sia nelle cellule vegetali che animali.
3) Infine la formazione dei recettori nucleari della T3 (TH-Rs = geni e relative proteine), che hanno permesso un migliore adattamento dei vertebrati all’ambiente terrestre. Infatti, negli anfibi, tramite la metamorfosi, le branchie si sono lentamente trasformate in polmoni e le pinne in arti. La maggiore gravità terrestre ha stimolato inoltre la ossificazione degli arti e dello scheletro. Mentre le maggiori escursioni termiche terrestri hanno sviluppando l’azione calorigena della T3, come protezione degli animali terrestri più evoluti: uccelli e mammiferi. E’ importante qui ricordare che i TH-Rs geni sono anche c-erbA oncogeni, che sono implicati come geni onco-soppressori in diversi tumori umani non-tiroidei, in particolare gastrici e mammari (Wang et al., 2002; Li et al. 2002). Nel 2001, Hays ha riportato sulla rivista statunitense “Thyroid” che “ è sorprendente che il contenuto totale dello iodio nel corpo umano sia ancora oggi incerto, e che dopo molti anni di ricerche, il metabolismo cellulare dello iodio ed il pool dello iodio extra-tiroideo siano ancora materia di speculazione e così pure la composizione chimica dello iodio extratiroideo sia ancora sconosciuta”. Questa affermazione fa risaltare le ricerche di Gribble (1996) e di Dembitsky e Tolstikov (2003) che hanno recentemente descritto più di 110 composti iodati presenti in organismi viventi animali e vegetali. Le due maggiori specie chimiche dello iodio presenti nell’interno delle cellule hanno differenti proprietà fisico-chimiche: lo iodio molecolare (I2) è idrofobo ed è capace di iodinare i doppi legami degli acidi grassi poli-insaturi delle membrane cellulari (PUFA), formando iodio-lipidi. Invece l’acido ipoiodico (HOI) è idrofilo e solubile nell’acqua. Recentemente Aceves e coll. (2005) hanno riportato, per la prima volta, che la percentuale di radio-iodio presente negli omogenati di tessuto mammario è del 40 % nella frazione lipidica, del 50 % nella frazione proteica e del 8 % nella frazione nucleare; e che negli omogenati di tessuto mammario di ratte la somministrazione di ioduri diminuisce significativamente la perossidazione lipidica. I pesci marini (come i selaci) sono ricchi di iodio e hanno anche meno tumori dei pesci di acqua dolce. Il 7 ottobre 1999 il Comitato del Senato USA ha ufficialmente dichiarato: “ Il Comitato ha notato la inusuale bassa incidenza di cancro in pesci marini (ricchi di iodio: NdT) come squali e razze , per cui incoraggia ricerche sul sistema immunitario di questi pesci per individuare sostanze anti-tumorali attive anche nell’ uomo”.
Fig. 5. Scintigrafie con I-123 total body sequenziali umane. La I-captazione in tutti i tessuti captanti è mediata dal NIS delle membrane cellulari. Nelle scintigrafie si notano oltre alla tiroide, altri tessuti iodiocaptanti: alla ventesima ora circa il 70 % dello radioiodio iniettato in vena è presente in sede extratiroidea: nella mucosa gastrica, epidermide, plessi coroidei cerebrali, ghiandole salivari ed inoltre, qui non visibili, nel timo fetale e nelle ghiandole mammarie (solo in gravidanza ed allattamento) ( Cortesia del Dr. G. Boni dell’Università di Pisa).
Nelle scintigrafie corporee con radio-iodio si notano oltre alla tiroide, altri tessuti iodiocaptanti: alla ventesima ora circa il 70 % dello radioiodio iniettato è presente in sede extratiroidea: nella mucosa gastrica, epidermide, plessi coroidei cerebrali, ghiandole salivari ed inoltre, qui non visibili, nel timo fetale e nelle ghiandole mammarie ( captanti solo in gravidanza ed allattamento). La tiroide capta in modo progressivo, mentre gli altri organi hanno un rapido accumulo ed una rapida dismissione del radio-iodio. La I-captazione è dovuta alla azione dei rispettivi simporter dello ioduro (NIS), che pur essendo simili, nella tiroide è filogeneticamente ed embriologicamente più evoluto, infatti è più affine per lo ioduro e risponde allo stimolo del più “moderno” TSH. Solo la tiroide, però, possiede il follicolo tiroideo che gli consente l’accumulo e il deposito di iodio-composti (TG). La iodiocaptazione tiroidea nel feto umano è infatti presente solo dalla 12° settimana di vita fetale e la formazione filogenetica della tiroide è anch’essa relativamente recente, risalendo a solo 400-500 milioni di anni fa’, quando i primi vertebrati marini cominciarono a popolare le acque degli estuari e poi dei fiumi terrestri carenti di iodio. A tale epoca della evoluzione i pesci di acqua dolce hanno cominciato ad utilizzare la vitamina C (acido ascorbico), che i vegetali avevano probabilmente allora iniziato ad produrre a scopo antiossidante. Infatti i pesci d’acqua dolce soffrono di “scorbuto” e di anomalie vertebrali causate da carenza di vitamina C, le quali regrediscono se tali pesci vengono rimessi in acque marine. In ambiente marino tali pesci possono utilizzare antiossidanti marini più primitivi, che sono in grado compensare il deficit alimentare di vitamina C. Infatti molti biologi ora ritengono che gran parte dei vertebrati si siano sviluppati morfologicamente e metabolicamente proprio nelle acque degli estuari (Purves e coll, 1998).
Fig. 6 . Salmoni coltivati in acqua dolce che mostrano sintomi di “scorbuto” ed anomalie della colonna vertebrale (scoliosi e lordosi) causate da carenza di vitamina C.
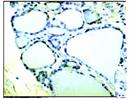
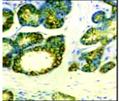
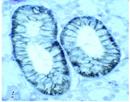
Fig. 7. Espressione del NIS, evidenziata da anticorpi colorati anti-NIS, nelle membrane plasmatiche in sede baso- laterale di: A: Tiroide; B: Mucosa gastrica; C: Mammella in allattamento; D: Ghiandola salivare ( Wapnir, 2003 ).
Storia evolutiva dello iodio negli esseri viventi: Dai più antichi: le alghe, gli invertebrati, i pesci, gli anfibi, i rettili, ai più recenti: i mammiferi fino all’ Homo Sapiens moderno.
Le alghe captano e trattengono gli ioduri in modo omogeneo e diffuso, le Laminarie contengono circa 1-3 % del peso secco di iodio. Le alghe marine (alghe verdi-azzurre) furono i primi esseri viventi a produrre ossigeno ( più di 3 miliardi di anni fa’), per cui hanno utilizzato efficaci e reperibili sistemi antiossidanti tra cui gli ioduri e il selenio (Pedersén e coll. e Kuepper e coll). Le alghe producono circa 80 % dell’ossigeno atmosferico, e costituiscono il primo anello della catena alimentare nutrizionale marina che trasferisce iodio, selenio e acidi grassi n-3 ai pesci e agli animali. Le acque salso-bromo-iodiche delle sorgenti termali del nostro entroterra (Salsomaggiore, Abano, Castrocaro ecc.) derivano da enormi praterie di alghe che durante la formazione della penisola italiana (circa 20-30 milioni di anni fa’) sono state ricoperte e sollevate da altri strati geologici, mantenendone però gli elementi algali primitivi mineralizzati come lo iodio, il bromo, il sale, il metano ecc. Risalendo la scala evolutiva filogenetica, possiamo vedere, circa 800 milioni di anni fa’, la comparsa di una notevole iodocaptazione nell’esoscheletro della conchiglia, e anche delle spugne e dei coralli (Brown-Grant, 1961). Roche (1951) aveva dimostrato negli invertebrati la presenza di MIT, DIT e T4, che vengono secreti ed espulsi dalle cellule nell’esoscheletro, dopo aver ceduto l’elettrone dello ioduro. Circa 500 milioni di anni fa’ nei pesci marini è comparsa una iodiocaptazione generalizzata, con minimo accumulo tiroideo, poco necessario per la facile reperibilità di ioduri nel mare. I follicoli, qui non ancora “tiroidei”, sono privi di capsula e si presentano scarsi e diffusi negli organi interni addominali. I pesci marini contengono alte quantità di iodio in gran parte inorganico circa 500-800 microgrammi (mg) per kg. Alcuni pesci marini migratori (anadromi) come le lamprede (Youson et al.,1997) , i salmonidi ecc. risalgono dal mare i fiumi fino alle sorgenti, dove muoiono inspiegabilmente dopo essersi riprodotti. In questo modo tali pesci marini riportano nei territori I-carenti dell’interno notevoli quantità di iodio e di selenio ( e anche di omega-3), consentendo la vita ed il benessere di altre specie animali tra cui gli uomini che, tramite loro, assimilano tali essenziali oligoelementi. Circa 400 milioni di anni fa’ quando, per la ricerca di cibo o per sfuggire ai predatori, alcune specie di pesci cominciano a lasciare il mare per abitare le acque dolci e I-carenti dei fiumi terrestri iniziano anche le malattie da carenza di iodio e di altre sostanze di origine marina. Nei pesci d’acqua dolce (in cui lo iodio è carente) i follicoli tiroidei sono più numerosi ed aggregati spesso tra le branchie. I loro tessuti contengono molto meno iodio rispetto ai pesci marini (circa 20 mg/ kg) ed è in gran parte iodio organico ( MIT, DIT , T4 ). I pesci d’acqua dolce I-carenti presentano inoltre anche difetti della immunità e maggiori malattie infettive, parassitarie, arteriosclerotiche e tumorali dei pesci marini (selaci). Infatti i selaci e i pesci marini in genere soffrono molto più raramente di neoplasie. L’alimentazione con pesce di mare è inoltre utile nella prevenzione di alcune importanti patologie tumorali, cardiache ed arteriosclerotiche nell’uomo.
Fig. 8. Metamorfosi della rana da animale acquatico (il girino) a rana terrestre. La quantità ambientale di iodio, necessario a formare la tiroxina endogena, innesca il meccanismo della metamorfosi. E’ evidente la “spettacolare” apoptosi (morte programmata) della coda, delle pinne, delle branchie che si trasformano negli arti ed anche dello stomaco, che da stomaco primitivo erbivoro si trasforma in stomaco pepsino-acido secernente proprio dei mammiferi carnivori. (Circa 300-400 milioni di anni fa’) (Ishizuya-Oka et al., 2003)
Lo studio della azione dello iodio nello sviluppo e metamorfosi degli anfibi è importante e riassume la strategia evolutiva di adattamento degli animali acquatici (pesci) in animali terrestri. Gli anfibi hanno una vita larvale nell’acqua dolce ( non esistono anfibi marini !) ed una vita adulta “terrestre” da circa 370 milioni di anni fa’. E’ esclusivamente la quantità ambientale di iodio, che consente la formazione della tiroxina endogena, ad innescare il meccanismo della metamorfosi, adattando così gli anfibi adulti alla vita “terrestre”con la formazione di polmoni, di epidermide lubrificata, arti deambulanti ossificati, di idonea circolazione cardio-polmonare ed anche di mucosa gastrica acido-pepsino-secernente, simile a quella dei mammiferi. Il girino carente di iodio non riesce a metamorfosare e muore presto come girino. C’è stata anche una evoluzione “morfologica” macroscopica dei follicoli della ghiandola tiroidea, i quali durante l’evoluzione (dai pesci, anfibi, rettili, uccelli e mammiferi) sono diventati sempre più definiti, incapsulati ed organizzati nella tiroide in sede pre-tracheale.
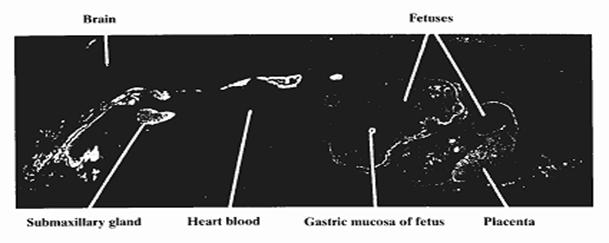
Fig. 9. Distribuzione dello iodio-125 (in bianco) nella autoradiografia di una topolina gravida dopo 1 ora dalla iniezione endovenosa dell0 I-125. E’ evidente la alta I-captazione nella mucosa orale e nelle ghiandole salivari, nella placenta e nelle mucose gastriche dei feti. (Autoradiografie di Ullberg ed Ewaldsson, 1964. Riprodotte per cortesia di Acta Radiologica)
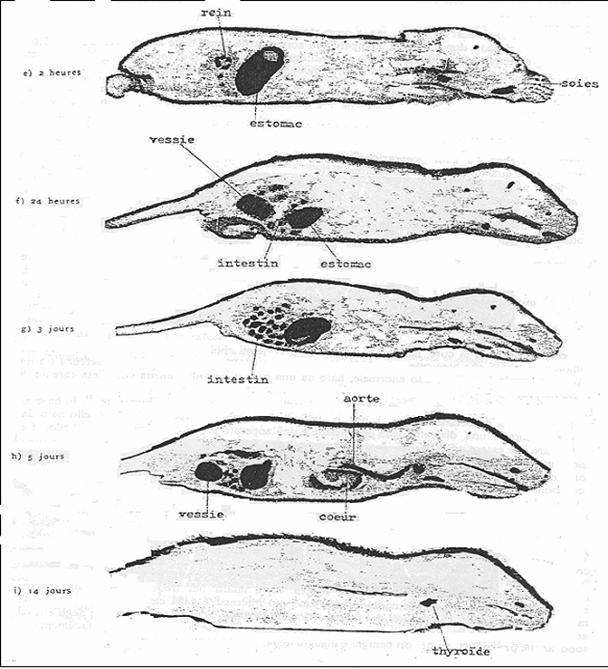
Lo studio autoradiografico di Ullberg ed Ewaldsson (sopra riportato) è importante in quanto studia il comportamento dello iodio e del NIS anche durante la gravidanza, e nella mammella ed in particolare nei tessuti fetali dei mammiferi, la cui origine risale a circa 200 milioni di anni fa’. Dopo un solo minuto dall’iniezione endovenosa dello I-131, si evidenzia la precocissima e rilevante captazione degli epiteli orale, salivare e gastrico e successivamente anche della ghiandola mammaria, della topolina gravida. La I-captazione della mucosa gastrica è molto alta, e dalla mucosa gastrica lo iodio viene secreto nel succo gastrico e riversato nell’intestino, dove viene riassorbito e ricaptato dallo stomaco, oltre che dalla tiroide, creando così una circolazione (NIS mediata) salivare e gastro-enterica-tiroidea dello iodio, che si perpetua fino ad eliminazione completa per via reno-vescicale ( Hays et al, 1965; Josefsson e coll., 2006). Quindi lo stomaco, più della tiroide, sembra avere un primitivo ruolo centrale nel metabolismo dello iodio su scala evolutiva. Dopo 5 giorni lo radioiodio è ancora ben visibile nelle pareti della aorta, e dopo 14 giorni è visibile solamente nella tiroide, nell’aorta e nella pelliccia dei ratti. In queste sedi lo iodio è presente sotto forma di iodo-composti, probabilmente proteici e lipidici, tuttora chimicamente non identificati.
FIG. 10. Autoradiografie con I-131 sequenziali nel ratto. La autoradiografia con I-131 nel ratto mostra la cospicua I-captazione della parete arteriosa della aorta, ben evidente anche dopo 5-14 giorni dalla iniezione del radioiodio, che potrebbe chiarire la sede della azione antiossidante ed anti-aterosclerotica dello ioduro. ( Da Pellerin, 1961; Riprodotte per cortesia di Path. Biol.)
Le autoradiografie con radio-iodio evidenziano che della mucosa gastrica la parte I-captante è costituita solo dalle cellule muco-secernenti della superficie e dei colletti delle ghiandole gastriche, che costituiscono proprio quelle foveole gastriche da cui si originano i carcinomi gastrici. Evans e coll. hanno dimostrato in ratte tiroidectomizzate che la tiroxina (T4) è molte volte piu’ efficace dello iodio inorganico (iniettato sottocute) nel ripristinare la normalità in molteplici funzioni fisiologiche, come la funzione ovarica e la crescita corporea, il metabolismo, il ritmo cardiaco, e le funzioni riproduttive, surrenali, timiche e ipofisarie. Inoltre Goethe e coll. hanno dimostrato che i recettori ormonali nucleari della T3 non sono indispensabili per la vita nelle cavie. Secondo i fisiologi, al contrario dello iodio in sé, la ghiandola tiroidea, anche se importante, non è indispensabile per la vita e gli effetti della sua asportazione si manifestano tardivamente, solo dopo 2-3 settimane. Mentre lo iodio e la T3 sono capaci di trasformare il girino acquatico in una rana terrestre strutturalmente “più evoluta”, la tiroidectomia e l’ipotiroidismo nei mammiferi sembrano costituire (al contrario della azione pro-evolutiva della metamorfosi) una sorta di “rettilizzazione”, cioè quasi una regressione filogenetica allo stadio precedente di rettile, di cui vengono riacquistate alcune caratteristiche fisiche e metaboliche come: la pelle ispessita, secca, squamosa e con perdita di peli, e la digestione, i riflessi, il battito cardiaco rallentati, con riduzione di tutto il metabolismo, accumulo di lipidi, ipotermia ed infine iperuricemia metabolica (Venturi, 2000). Nei primati e nei uomini la tiroide è ben organizzata e a forma di farfalla, in sede pre-tracheale, dove, come un rudimentale termostato, può meglio avvertire e rispondere alle variazioni termiche ambientali. Negli uomini affetti da cretinismo endemico da carenza iodica sono evidenti i danni fisici, neurologici, mentali, immunitari (Marani e Venturi, 1985) e riproduttivi. Nel 1998 su Geographical Review, Dobson ha ipotizzato che la scomparsa dell’uomo di Neanderthal avvenuta circa 35.000 anni fa’, sia stata favorita dalle maggiori capacità fisiche, intellettive e di adattamento dell’ homo sapiens moderno, dovute al maggiore intake iodico. Ciò grazie anche al miglioramento dietetico e genetico del NIS, divenuto più efficiente nel captare lo iodio. Infatti, negli scheletri dell’uomo di Neanderthal, Dobson ha rilevato le stigmate ossee del cretinismo endemico, con arti corti e tozzi e microcefalia. A differenza del Neanderthal, l’homo sapiens moderno aveva habitat più vicino ai mari e dieta ricca di pesce marino ricco di iodio. Broadhurst (2002) e Cunnane (2005) hanno recentemente riportato che lo iodio è stato l’elemento più importante insieme agli acidi grassi poli-insaturi (omega-3) nel favorire il processo di evoluzione del cervello umano e del conseguente sviluppo dell’intelligenza umana, permettendone così un migliore adattamento ambientale.
FIG. 11 Mappa mondiale delle aree di endemia di gozzo da carenza iodica (ombreggiate obliquamente) spesso circostanti a catene montuose (in blu), prima della effettuazione della iodio-profilassi nel mondo (da Kelly e Snedden, OMS, 1960).
Fig. 12. Mappa mondiale delle Nazioni riguardo alla attuale nutrizione iodica ( Da ICCIDD - OMS, 2003) Ancora oggi secondo l’OMS, più di tre miliardi di persone sulla terra vivono in aree distanti dal mare e con carenza ambientale di iodio e soffrono di tireopatie, di danni neurologici e somatici ed inoltre di diminuzione delle difese immunitarie e di ridotta fertilità con danni alla prole; inoltre tali persone sembrano più soggette a patologie tumorali della tiroide, dello stomaco, della mammella e alle patologie delle ghiandole salivari e della bocca con anche una maggiore perdita dei denti (Venturi e al. 2000, 2001; Aceves e al.,2006; Szybinski e al., 2004; Abnet e al., 2005.a, 2005.b, 2006). La supplementazione iodica, con sale da cucina iodato, con foraggio o con concime composto di alghe marine ricche di iodio, è in grado di prevenire questi danni negli animali ed anche nei vegetali per quanto riguarda la loro suscettibilità a infezioni microbiche e parassitarie aumentandone i meccanismi di difesa antiossidante (Saker e al. 2001; Fike e al., 2001; Cabello e al., 2003; Food and Nutrition Board of USA, 2001). E’ ipotizzabile che nella vasta gamma degli antiossidanti naturali vi sia una sorta di “gerarchia filogenetica”, in cui gli antiossidanti primitivi marini come lo iodio, il selenio ecc. svolgano un ruolo più importante nella vita riproduttiva e nello sviluppo fetale di quelli che si sono evoluti più recentemente ( ad esempio il licopene, le antocianine, il resveratrolo, molti polifenoli ecc.), come hanno riportato a proposito dello iodio Dunn e Delange (2001). Nei territori I-carenti invece tali patologie coesistono sia negli uomini che negli animali, in particolare negli erbivori. Tutto ciò fa’ supporre che il processo di adattamento evolutivo dei vertebrati terrestri alla carenza iodica ambientale non sia ancora terminato. Pubblicato in: “Guida alla Epidemiologia e Prevenzione del Gozzo Endemico” di D. Meringolo, D. Bianchi e B. Bellanova. 8 Luglio 2006, Bologna. Pagg. 11-25 Atti del Convegno Nazionale di Endocrinologa Clinica. 8 Luglio 2006 – Bentivoglio (BO) |
||||
![[Bohr Model of Iodine]](iodio-antiossidanti-evoluzione_file/image002.jpg)


 A
B
C D
A
B
C D


|
02-gen-2008 Scopri come dimagrire con il riso! E' facile e non richiede sforzi!
Vuoi una dieta personalizzata gratis? Cerchi consigli per dimagrire? Scrivimi i tuoi obiettivi e le tue abitudini alimentari ed io risponderò.
Consigli su quali e quanti esercizi fare? Vuoi capire come si effettua un allenamento completo? Entra.
La frutta e le sue proprietà
I principali alimenti che compongono il modello mediterraneo.
Vuoi comparire su internet e gareggiare virtualmente con altri Bodybuilder? Entra.
E' importante riposarsi correttamente?
Scopri come funziona il sistema immunitario e quali alimenti e integratori assumere per aiutarlo!
MANTENERE UNA DIETA EQUILIBRATA ED UNO STILE DI VITA SALUTARE AUMENTA LE ASPETTATIVE DI VITA E LA SUA QUALITA'. QUINDI SII ATTENTO A COSA MANGI E PRATICA SPORT CON PASSIONE! DIMAGRIRE E DIVENTARE MAGRI IN MODO SANO PUO' ESSERE MOLTO SEMPLICE! Englih Version New
|