|
BIOCHIMICA DELLA BIOLUMINESCENZA
Alcune molecole possono essere trattate in modo da
provocare l'emissione di luce; questo fenomeno viene chiamato chemioluminescenza.
Questi sistemi modello dimostrano che una molecola emettente luce
deve essere portata a uno stato di eccitazione elettronica con un
alto potenziale di energia; parte della energia di eccitazione viene
poi liberata sotto forma di un fotone mentre la molecola ritorna
allo stato nativo
A => eccitamento => A* => A + hv
La bioluminescenza si basa sullo stesso principio.
Gli studi biochimici di sistemi privi di cellule appartenenti a
diversi organismi hanno dimostrato che i meccanismi di emissione
di luce sono estremamente vari; è probabile che questa particolare
proprietà funzionale si sia sviluppata indipendentemente
nei diversi gruppi biologici. Nei batteri la bioluminescenza si
associa a un consumo di O2; pertanto, l'emissione di luce avviene
soltanto in condizioni di aerobiosi.
L'emissione di luce da parte di una sospensione di batteri luminescenti
costituisce uno dei metodi più sensibili per la rivelazione
di tracce di ossigeno disciolto.
La reazione per la bioluminescenza ,catalizzata dall'enzima luciferasi,
avviene attraverso la ossidazione del FMNH2 e di un'aldeide alifatica
da parte dell'ossigeno molecolare
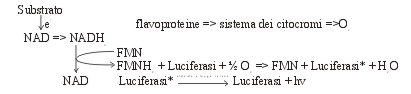
La luminescenza batterica si può considerare
come una naturale deviazione della normale catena respiratoria batterica,
per mezzo quale una frazione di elettroni (e-) viene trasferita
dal substrato all'ossigeno.
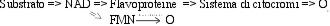
Il legame fisiologico tra le due catene viene indicato
dall'effetto del cianuro. Questo inibitore del trasporto di elettroni
via citocromi, aggiunto a una sospensione batterica bioluminescenti
causa un aumento della bioluminescenza. Quando gli elettroni giungono
all'ossigeno passando attraverso la deviazione, non si forma ATP,
perché la maggior parte dell'energia viene emessa sotto forma
di luce.
L'enzima luciferasi mostra un tipo unico di sistema di regolazione
chiamato autoinduzione. I batteri luminescenti producono una sostanza
specifica, l'autoinduttore, il quale viene accumulato nella coltura
durante la crescita, e quando la quantità di questa sostanza
ha raggiunto un livello critico, avviene l'autoinduzione dell'enzima.
Così le colture di batteri luminescenti a bassa densità
non sono luminose, ma incominciano a diventarlo quando la crescita
raggiunge una densità sufficientemente alta così che
l'autoinduttore può accumularsi e funzionare. Sebbene l'autoinduzione
avvenga in tutte le specie di batteri luminescenti, l'autoinduttore
di una specie è specifico per essa e non indurrà l'induzione
di un enzima in un'altra specie. A causa del fenomeno dell'autoinduzione
è ovvio che un singolo batterio libero nel mare non è
luminoso, perché l'autoinduttore non si può accumulare.
Il motivo della dipendenza per la luminescenza dalla densità
è chiaro per quelle colonie di batteri luminescenti in simbiosi,
perché la luminescenza si sviluppa quando si è raggiunto
unalta densità nell'organo in cui avviene la simbiosi (fotoforo).
Ma è meno chiaro perché la luminescenza dipenda dalla
densità, per quei batteri che vivono liberi nell'ambiente
marino. Un idea è che questi batteri sono luminescenti solo
quando crescono saprofiticamente su pesci in decomposizione, dove
possono crescere con alte densità cellulari formando colonie
luminescenti. Questa luminescenza servirebbe per attrarre un maggior
numero di organismi per ingerire il materiale, così da essere
sicuri del trasferimento dei batteri ad un altro habitat favorevole
per la crescita.
L'energia usata per la luminescenza è considerevole: sotto
condizioni di massima luminescenza circa il 10 - 20 % dell'O2 cellulare
totale viene impiegato per il sistema della luciferasi.
 |
| |
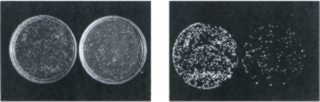
| Fig 3.1 Piastre di Photobacterium
in luce e al buio. I fotobatteri, per la bioluminescenza, necessitano
di O2 e di un aldeide a lunga catena che favorisce di 1000 volte
il processo di liberazione del fotone durante il ritorno allo
stato originario della luciferasi eccitata. |
|

