
Struttura secondaria del DNA
La struttura secondaria del DNA è la ben conosciuta doppia elica. Fu dedotta dall'analisi degli spettri di diffrazione di fibre di DNA Gli spettri di diffrazione delle fibre non contenevano le informazioni molecolari trovate negli spettri di diffrazione del singolo crisstallo, ciononostante sia gli spettri di diffrazione che nle proprietà delle fibre fornirono informazioni essenziali per dedurre la struttura della doppia elica
-
Gli spettri mostravano la presenza di un'elica
-
la spaziatura tra le macchie mostravano che c'erano 10 basi per giro d'elica
-
la densità delle fibre indicava la presenza di due molecole per elica.
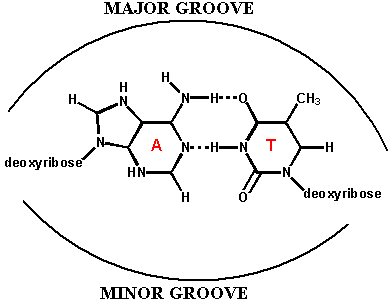
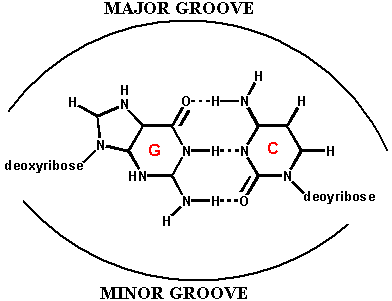
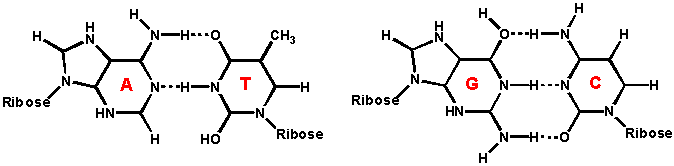
Watson e Crick proposero che la doppia elica fosse stabilizzata dalla formazione di legami ad idrogeno tra le basi presenti sui filamenti opposti secondo lo schema A-T e G-C
Secondo il loro modello, lo scheletro idrofilo costituito dalla sequenza zucchero-fosfato si trovavano all'esterno a contatto con l'acqua, mentre le basi di carattere idrofobico erano impacchettate perpendicolarmente all'asse dell'elica nell'interno.
I due filamenti di DNA sono complementari e si sviluppano in direzione opposta, sono perciò antiparalleli.
Benchè le basi si trovino all'interno, esse possono interagire con altre molecole attraverso le due spirali profonde chiamate solco monore e solco maggiore. Questo modello teneva conto del fatto che nella composizione delle basi del DNA si verificava che in percentuale A=T e G=C.
Stabilità della doppia elica
La formazione della doppia elica è favorita da
- Interazioni idrofobiche derivanti dall'impaccamento delle basi
- dalla formazione di legami ad Idrogeno tra le coppie di basi.
|
La formazione della doppia elica è sfavorita da
- Repulsione elettrostatica dei fosfati che si trovano sullo scheletro dei filamenti
- Dall'entropia conformazionale
- Formazione di legami ad Idrogeno con l'acqua.
Le Possibili strutture della doppia elica
Il modello di doppia elica proposto da Watson e Crick rappresenta la struttura del BDNA. La struttura ai raggi X confermò essenzialmente il modello proposto, ma mostrava come essa fosse in realtà una semplificazione in quanto
- la struttura molecolare mostrava molte variazioni e distorsioni locali rispetto alla struttura ideale
- E' importante comprendere che la struttura secondaria del DNA non è rigida ma flessibile e dipende dalla esatta sequenza dei nucleotidi e può essere modificata in seguito ad interazioni con proteine ed altre molecole.
Sotto altre condizioni è possibile che si produca un altro modello di
elica ADNA.
Recentemente è stata descritta una ZDNA,
destrosa, il cui significato biologico è sconosciuto. Qui c'è un confronto tra i diversi modelli di elica. La forma A è
la conformazione ad elica adottata dal DNA e RNA, ARNA,
in quanto l'ossidrile in posizione 2' del ribosio impedisce stericamente la
formazione della conformazione B.
 La maggior parte el DNA della maggior parte dei microrganismi è
una doppia elica nella sua forma B
La maggior parte el DNA della maggior parte dei microrganismi è
una doppia elica nella sua forma B
La struttura molecolare suggerisce che la forma B è quella predominante
in natura poichè essa, a differenza della A, può inserire
una spina di molecole d'acqua (in verde) nel solco minore.
I legami ad Idrogeno apportati dall'acqua aiutano a stabilizzare l'elica
nella forma B
In alcuni microrganismi il DNA è circolare mentre in altri è
lineare
Il DNA delle cellule procariote è immenso.
Il DNA di una Drosophila melanogaster (mosca della frutta)
ha un peso molecolare di 4 x 1010 g/mol (~6.5 x 107 coppie di basi) e sarebbe lunga 2 cm se fosse estesa.
Il DNA complessivo di tutte le cellule di un uomo si estenderebbero
per 50 volte la circonferenza del sole.
Dal momento che eccedono le dimensioni della cellula deve trovarsi
sotto forma estremamente condensata.
La Replicazione semiconservativa del DNA
Denaturazione e Rinaturazione del DNA
Benchè la doppia elica sia relativamente stabile nelle condizioni fisiologiche,
è possibile che la denaturazione della struttura secondaria possa verificarsi
in seguito a vari fattori:
Enzimi quali RNA polimerasi ed elicasi
Valori elevati di pH che portano a repuilsioni elettrostatiche tra i gruppi
fosfato carichi negativamente.Tali cariche possono essere neutralizzate da
ioni o proteine ed un loro cambiamento di concentrazione può condurre
a separaioni nella catena.
Sostanze chimiche possono abbassare l'energia di impaccamento delle basi
nella doppia elica
Un altro fattore che controlla la denaturazione è la temperatura
in quanto l'entropia dello stato denaturato ( random coil) è maggiore
dello stato elicoidale.
Infatti l'enegia libera per la transizione Elica-->random coil è DG = DH - TDS.
Per questa transizione il DH eletrostatico<0
e DSconfigurazione>0: entrambi favoriscono il processo. Perciò perchè la doppia elica sia stabile sotto queste condizioni è necessario che si formi un DH>0 nella transizone elica-> random coil che favorisca lo stato elicoidale. Questo DH positivo si origina dalla rottura dei legami di Vander Walls dell'impaccamento delle basi che rappresenta il fattore principale che favorisce la formazione dell'elica.
Alla temperatura fisiologica il DH deve essere maggiore del DHelett-TDSconfigurazione perchè l'elica sia possibile.
A temperature maggiori prevale naturalmente il fattore entropico e quindi il segno del DG cambia da negativo a positivo
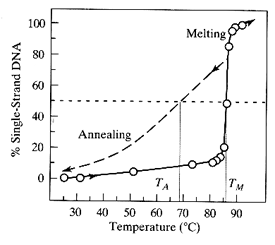 La denaturazione può essere monitorata a 260nm zona d'assorbanza delle basi la cui assorbanza diminuisce quando sono impaccate rispetto alla randomcoil.
La denaturazione può essere monitorata a 260nm zona d'assorbanza delle basi la cui assorbanza diminuisce quando sono impaccate rispetto alla randomcoil.
La temperatura di transizione, Tm, è alquanto ripida poichè la denaturazione è un fenomeno fortemente cooperativo.
Si noti che la curva di riformazione della doppia elica è differente da quella di fusione e che la temperatura di riassociazione Ta è minore della Tm. (isteresi)
La ripidezza della curva di fusione suggerisce che il processo sia altamente cooperativo.
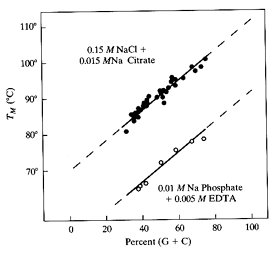 La Tm dipende sia dal rapporto (G+C)/(A+T) che dal contenuto salino della soluzione. Si rammenti che la coppia GC è stabilizzata da 3 legami ad H mentre la coppia AAT da due.
La Tm dipende sia dal rapporto (G+C)/(A+T) che dal contenuto salino della soluzione. Si rammenti che la coppia GC è stabilizzata da 3 legami ad H mentre la coppia AAT da due.
l'energia dell'impacchettamento delle basi tra un GC e un GC o AT è più forte di quello delle coppie AT.
Di conbseguenza un più alto contenuto in GC porta ad una migliore stabilità dell'elica sia per un maggior numero di legami ad H che per interazioni di Vander walls.
Una forza ionica alta scherma le repulsioni elettrostatiche dei gruppi fostato nello scheletro
Molecole che si intercalano tra le basi nel DNA causano un salto di basi nella trascrizione del DNA. (etidiobromuro). Le aflotossine che si ritrovano nelle granaglie e noci ammuffite sono agenti alchilanti che modificano la guanina portando così a mutazioni prive di senso.
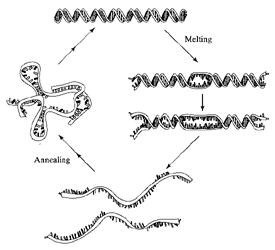 Modelli di fusione e di riassociazione
Modelli di fusione e di riassociazione
La fusione inizia alle estremità o nelle zone di più alto
contenuto di coppie AT. All'aumento della temperatura le regioni in cui
si è avuto la fusione si estendono fino alla separazione completa
dei due filamenti. La riassociazione è un processo bimolecolare
ma ci sono processi in competizione in cui possono aversi accoppiamenti
di base sia in un filamento che tra filamenti, portando alla fine ad una
doppia elica stabile.