|
Citologia |
| Membrana plasmatica | Reticolo endoplasmatico | Apparato del Golgi | Centrioli | Nucleo |
| Citoscheletro | Mitocondri | Ribosomi | Cloroplasti | Citoplasma |
| Cromosomi | Lisosomi e Perossisomi | Parete Cellulare |
Morfologia cellulare
|
Reticolo endoplasmatico
col nome di
reticolo endoplasmatico
viene designato
un sistema di cisterne e di canalicoli
delimitati da membrane di natura lipoproteica, costituito da due
compartimenti
che comunicano tra loro ma che differiscono
per costituzione e funzioni
svolte:
·
REG
- reticolo endoplasmatico
granulare (o
rugoso RER): strutturato a
cisterne
·
REL
- reticolo endoplasmatico
liscio:
strutturato a canalicoli
Questi due
compartimenti intercomunicanti hanno in comune un costituente di membrana
dello spessore di 5¸6 nm, la cui struttura è simile a quella di altre
citomembrane. I compartimenti differiscono per la forma delle cisterne
delimitate da queste membrane e, nel caso del REG, per la presenza sulla
faccia esterna di granulazioni dette
ribosomi,
che vi sono ancorate. Struttura laminare del REG
Gli elementi
caratteristici del REG
consistono in sottili
lamine
composte da due membrane accollate e delimitanti una cavità a forma di
sacco appiattito, più o meno allungato, le cui membrane sono ricoperte di
ribosomi. Disposizione e numero dei sacculi variano in rapporto al tipo cellulare e all’intensità funzionale del momento. Quando il REG è molto sviluppato, la disposizione dei sacculi segue un determinato schema, essendo fra loro paralleli e dislocati in determinati distretti della cellula. Nelle cellule ghiandolari pancreatiche e salivari, tutti i sacculi occupano la porzione basale del citoplasma; la loro quantità è così elevata che in microscopia ottica danno a quest’area cellulare un aspetto pieghettato. Negli epatociti i sacculi sono raggruppati a formare i corpi di Berg e hanno spesso una disposizione concentrica. Nelle cellule nervose multipolari delle corna anteriori del midollo spinale i sacculi si riuniscono in piccoli territori, che corrispondono ai corpi di Nissl della microscopia ottica.
Nei plasmociti
occupano tutto il citoplasma: la loro disposizione descrive curve
concentriche al nucleo. Anche le cellule meno attive contengono dei
sacculi, ma sono più radi e sono disseminati nel citoplasma in modo
disordinato.
La funzione fondamentale del REG è
legata al processo di secrezione
proteica
e consiste nel trasporto del prodotto che è stato sintetizzato a livello
dei ribosomi. L’associazione dei ribosomi alla membrana del reticolo
endoplasmatico consente alle proteine neosintetizzate di penetrare
all’interno delle cisterne in quanto, prima di essere liberate dalla
cellula, debbono subire una lunga serie di trasformazioni che possono
realizzarsi solo all’interno di un sistema chiuso. |
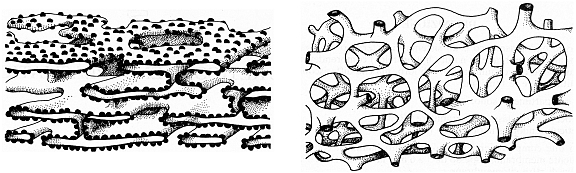
Fig. VI. 1 -
Rappresentazione schematica tridimensionale del reticolo endoplasmatico
granulare e liscio
Struttura tubulare del REL
Come abbiamo visto, il REG
ha struttura laminare, mentre il REL è costituito da un
labirinto di canalicoli
intercomunicanti
distribuiti in tutto il citoplasma,
senza ribosomi
adesi alla faccia esterna delle sue membrane. Ancora contrariamente al REG, il
REL ha la proprietà di
stabilire stretti contatti
coi mitocondri,
coi depositi di glicogeno e coi perossisomi. I microsomi
Le tecniche di
centrifugazione differenziale permettono di isolare le membrane del reticolo
endoplasmatico sotto forma di vescicole, i
microsomi,
ricoperte o meno di ribosomi. L’analisi biochimica di queste membrane, trattate
precedentemente con ribonucleasi per eliminare le ribonucleoproteine, rivela che
le membrane del reticolo endoplasmatico contengono:
o
proteine
strutturali e lipidi (30-50%),
o
enzimi
necessari alla sintesi proteica, al metabolismo dei lipidi, ai fenomeni di
detossicazione. Funzioni del reticolo endoplasmatico2.1. Funzioni di trasporto
Il reticolo endoplasmatico
assicura il trasporto dei seguenti elementi:
§
elettroliti
§
elettroni,
grazie a un sistema di trasferimento extramitocondriale
§
sostanze
elaborate nei vari distretti della cellula, convogliate nel mezzo extracellulare Funzioni di sintesia. Sintesi delle proteine
Il ruolo del
REG
nella sintesi proteica è stato analizzato precedentemente con le funzioni del
ribosoma. Le proteine sintetizzate sui ribosomi penetrano nel lume delle
cisterne del REG per mezzo di un trasferimento che avviene secondo un preciso
meccanismo: il ribosoma dà inizio alla sintesi del polipeptide associando tra
loro gli aminoacidi in una sequenza segnale (parte iniziale della catena
polipeptidica); la sequenza segnale interagisce con le proteine intramembranarie
del REG, dove le proteine intrinseche si avvicinano e si riuniscono in gruppo a
formare un tunnel; nella cavità delle cisterne del REG un’idrolasi rimuove la
sequenza segnale della proteina specifica; questo taglio segna la fine
della sintesi e l’inizio del trasferimento della molecola proteica neoformata
nel reticolo. I ribosomi che hanno ultimato la protidosintesi si staccano dalla
membrana e le loro subunità si separano. b. Sintesi dei lipidi
Il
REL
è responsabile della sintesi dei lipidi, in quanto le sue membrane sono dotate
di alcuni sistemi enzimatici responsabili dell’allungamento e della saturazione
degli acidi grassi. Il REL è molto abbondante nelle ghiandole sebacee, negli
spongiociti, nei luteociti, ovunque la sintesi dei lipidi sia molto attiva.
Interviene, per esempio, nella sintesi degli steroidi, che sono ormoni derivati
dal colesterolo. Il REL è capace di sintetizzare questa complessa molecola a
partire dall’acetato e la accumula nelle inclusioni lipidiche in attesa di
essere utilizzato nei mitocondri. Infine, nel REL vengono sintetizzati
fosfoIipidi e trigliceridi. c. Sintesi delle glicoproteine
Le proteine sintetizzate
dal REG possono associarsi con gli zuccheri, dando origine a glicoproteine
attraverso un processo di glicosilazione, che inizia nel REG e si completa
nell’apparato di Golgi. d. Sintesi delle membrane cellulari
Il sistema membranario
altamente differenziato del reticolo endoplasmatico, dove vengono sintetizzati
fosfolipidi e proteine, dà luogo alla formazione delle membrane cellulari. Lo
studio della citodifferenziazione indica che la comparsa del REG precede quella
del REL. Le proteine e i fosfolipidi vengono assemblati nel REG e poi
trasportati a costituire il REL. La quantità di fosfolipidi sintetizzati è
regolata dalla quantità di proteine capaci di legarsi ai fosfolipidi stessi, il
che significa che la quantità di nuove membrane sintetizzate è determinata dalla
quantità di proteine disponibili nel REG, dove si effettua l’associazione
proteine-fosfolipidi. e. Funzioni di immagazzinamento e di accumulo
Il reticolo endoplasmatico
immagazzina e concentra sostanze provenienti sia dall’ambiente extracellulare
che da quello intracellulare:
q
sostanze di origine
extracellulare:
i vacuoli di pinocitosi prelevano sostanze dall’ambiente extracellulare, per
esempio proteine, si portano nello ialoplasma e qui riversano il loro contenuto
nelle cisterne del REG
q
sostanze prodotte dalla
cellula: nell’epatocita
i granuli lipoproteici elaborati dalla cellula si concentrano, si accumulano e
vengono messi in riserva temporanea entro espansioni vescicolari del REL. Ma
l’esempio più sorprendente è certamente quello delle plasmacellule, che
producono gli anticorpi grazie a un REG molto sviluppato. Le plasmacellule di
animali iperimmunizzati contengono grandi quantità di anticorpi all’interno di
particolari vacuoli, i corpi di Russel, le cui membrane appartengono al
reticolo endoplasmatico. f. Funzioni di detossicazione
Le membrane del REL -
principalmente nel fegato e nel rene - trasformano molecole tossiche in composti
atossici prima di essere eliminati dall’organismo, frequentemente per intervento
del citocromo P 450. Per esempio, la detossicazione trasforma, per
idrossilazione, i composti tossici liposolubili in composti idrosolubili, che
vengono così eliminati dai reni.
esso presiede alla sintesi di enzimi e altre proteine, di grassi e sostanze fondamentali per la cellula, come alcuni ormoni steroidei
|