
SALIVAZIONE
La secrezione salivare é promossa da
tre paia di ghiandole principali: la parotide, la sottomascellare e la
sottolinguale, cui si aggiungono numerose ghiandole accessorie, o minori,
sparse in tutta la mucosa del cavo orale, tranne che in quella delle arcate
dentarie. La produzione salivare ammonta a circa 1000-1500 ml al giorno,
comprese le fasi digestive in cui è maggiore: nei periodi interdigestivi ne
viene prodotta circa 1 ml al minuto.
La secrezione salivare é composta di acqua, elettroliti, enzimi,
glicoproteine e fattori di crescita. La saliva è importante per lubrificare
ed ammorbidire il cibo al fine della deglutizione, per solubilizzare sostanze
che così possono essere gustate, per pulire la bocca e i denti prevenendo
così la carie, per dare inizio alla digestione degli amidi, per aiutare ad
allontanare dall'esofago le secrezioni del reflusso gastrico e per
l'articolazione della parola.
Anatomia funzionale
Le ghiandole salivari principali sono strutture tubulo-alveolari, che rappresenta uno schema della morfologia della
ghiandola sottomandibolare, mista.
L'aspetto acinoso è dovuto alle
estremità secretorie, che sono a loro volta classificate in tre gruppi
principali, a seconda del tipo di cellule che vi si trovano: sierose, mucose e
miste, che contengono elementi sia sierosi che mucosi. Le ghiandole salivari le
cui estremità secretorie contengono prevalentemente cellule sierose o mucose
sono classificate come ghiandole sierose o mucose, rispettivamente; quelle le
cui estremità secretorie contengono ambedue gli elementi sono considerate
miste. Le parotidi, che sboccano nel vestibolo orale con il dotto di Stenone
all'altezza del primo molare, sono ghiandole a secrezione in prevalenza sierosa,
che producono quindi un secreto molto fluido. Le sottolinguali e le
sottomandibolari, che sboccano sul pavimento della cavità orale, ai lati
dell'inserzione del frenulo linguale mediante il dotto di Wharton ed il dotto di
Rivino, sono di natura mista (la sottomandibolare prevalentemente sierosa,
quella sottolinguale prevalentemente mucosa).
Le cellule delle estremità secretorie sono caratterizzate da un nucleo situato
in posizione basale e dalla presenza del reticolo endoplasmico rugoso, che è
maggiormente sviluppato nelle cellule sierose. Ambedue i tipi di cellule
contengono granuli di secreto in posizione apicale che, nelle cellule sierose,
sono più piccoli e contengono amilasi insieme ad una quantità variabile di
glicoproteine. I granuli delle cellule mucose sono più grandi e ripieni di una
speciale glicoproteina, la mucina.
Il ramificato sistema di dotti che porta dalle estremità secretorie alla
cavità orale comprende i dotti intercalati, striati ed escretori (fig. 1). I
dotti intercalati, interposti fra le estremità secretorie ed i dotti striati,
sono tappezzati da piccole cellule cubiche con nucleo prominente e scarso
citoplasma. I dotti striati devono il loro nome alla apparente presenza di
striature basali nelle cellule epiteliali che li costituiscono, che altro non
sono se non vaste invaginazioni, organizzate in serie di colonne parallele,
della membrana cellulare intorno ai mitocondri. Queste invaginazioni potrebbero
essere importanti per aumentare la superficie lungo la quale avviene il
trasporto trans-epiteliale di elettroliti. I dotti striati sboccano in quelli
escretori, che si aprono nella cavità orale. Nella maggior parte delle
ghiandole salivari, l'epitelio dei dotti escretori é composto da due strati:
uno superficiale di cellule colonnari ed uno basale di cellule appiattite.
Le estremità secretorie ed i dotti intercalari sono circondati da cellule
mioepiteliali, in grado di comprimere le strutture sottostanti, forzando così
la saliva nei dotti principali.
Composizione della
secrezione salivare
Composti inorganici. La composizione della
saliva dipende dalla ghiandola che la produce, dallo stimolo che la provoca e
dalla velocità della produzione. La saliva possiede, infatti, l'importante
caratteristica di variare la propria composizione a seconda della velocità di
produzione: tanto più la secrezione salivare aumenta (come durante la fase
masticatoria), tanto più aumenta la componente idrica della saliva stessa, e ne
diminuisce la pressione osmotica, promuovendo una migliore diluizione degli
alimenti, se pure a scapito della quantità di principi attivi. I composti
inorganici maggiormente rappresentati sono sodio, potassio, cloro, bicarbonato,
calcio, magnesio, fosfato, fluoro e tiocianato.
A riposo, la saliva é ipotonica, con una concentrazione di sodio inferiore a 6
meq/l ed una di potassio intorno a 30 meq/l (figura 2). Sotto stimolazione, la
concentrazione del potassio diminuisce (pur restando maggiore di quella
plasmatica), mentre quella del sodio aumenta fino a poter raggiungere quella
plasmatica; anche le concentrazioni del bicarbonato e del cloro aumentano. Il pH
della secrezione salivare a riposo é inferiore a quello ematico, mentre, quando
il flusso salivare aumenta, può diventare leggermente superiore.
Il risultato netto dell'azione dei dotti é rappresentato dall'assorbimento di
NaCl e dalla secrezione di KHCO3, di cui il primo si verifica più velocemente.
Poiché l'epitelio dei dotti é relativamente impermeabile all'acqua, il liquido
intraluminale diventa ipotonico. Tanto la stimolazione parasimpatica quanto
quella adrenergica aumentano la produzione salivare anche inibendo il trasporto
di Na+ da parte dell'epitelio dei dotti.
Composti organici. I composti
organici sono costituiti prevalentemente da proteine e glicoproteine che vengono
sintetizzate, immagazzinate e secrete dalle cellule dei segmenti terminali. Fra
questi vi sono amilasi (ptialina), glicoproteine, frammenti secretori,
callicreina, lattoperossidasi e lattoferrina. Una piccola quantità di proteine
viene anche sintetizzata, immagazzinata e secreta dalle cellule dei dotti, come
il fattore di crescita neurale, il fattore di crescita dell'epidermide, il
lisozima e la ribonucleasi. Amilasi e glicoproteine costituiscono la più gran
parte della secrezione organica dei segmenti terminali. La glicoproteina secreta
dalla parotide lega il calcio ed é ricca di prolina. La proteina contiene circa
sei oligosaccaridi con tre residui di mannosio legati ad asparagina per
N-glicosilazione. La mucina, glicoproteina specializzata secreta dalle ghiandole
sottolinguali, é una proteina O-glicosilata con molti oligosaccaridi a catena
corta uniti da legami covalenti a residui di serina o di trionina.
La saliva contiene anche una certa quota di albumina, immunoglobuline,
tiocianato ed altre sostanze ad azione batteriostatica o addirittura
battericida, tali da contrastare la proliferazione batterica, quali
sialoperossidasi e lisozima. Ciò è importante in quanto la cavità orale é il
punto di ingresso degli alimenti, rappresentando il luogo del primo contatto tra
ambiente esterno ed interno (l'interno del canale alimentare é tutto da
considerarsi, infatti, ambiente esterno) perciò facilmente esposto ad infezioni
e sede di una abbondante flora batterica. In condizioni di ridotta secrezione
salivare, la cavità orale va facilmente incontro a fenomeni patologici quali
flogosi, fenomeni settici e facile insorgenza di carie. All'azione di protezione
chimica della saliva si associa quella fisica di contenimento della
proliferazione batterica mediante il continuo "lavaggio" determinato
dall'accoppiamento salivazione-deglutizione. Provvederà poi il pH gastrico ad
eliminare quasi completamente la presenza batterica nel bolo alimentare. In
condizioni normali, infatti, in duodeno si riscontrano valori minimi di
contenuto batterico, che ritorna su valori elevati solo nell'intestino crasso,
dove il tempo di permanenza del contenuto é maggiore (nell'ordine delle 24 ore)
rispetto a quanto si verifica nell'intestino tenue.
L'azione della ptialina, che scinde i carboidrati in frammenti più piccoli, si
limita essenzialmente ad un solo substrato: l'amido cotto (l'organismo umano non
dispone di alcun enzima per l'amido crudo e la cellulosa). Il tempo di azione
della ptialina é ridotto, ma supera quello della permanenza del bolo nella
cavità orale in quanto prosegue nello stomaco, fino alla inattivazione per
denaturazione operata dall'acido cloridrico. Lo stomaco é sprovvisto di enzimi
digestivi per i carboidrati ed inoltre il suo basso valore di pH impedirebbe
l'azione delle amilasi, che lavorano a valori di pH neutro o leggermente
alcalino.
Tuttavia il riempimento dello stomaco avviene per strati concentrici
(il cibo cade dall'esofago al centro dello stomaco e viene successivamente
rimescolato dai movimenti della parete gastrica: sono necessari circa 15-30
minuti, a seconda del contenuto, per ottenere un rimescolamento completo) e ciò
consente un certo margine temporale all'azione della amilasi salivare.
L'altra importante componente secretoria della saliva é la mucina (sostanza di
consistenza viscosa, compatta e filamentosa) che assolve a funzioni
lubrificanti. La mucina é fortemente adesiva ed impastando e rivestendo il bolo
alimentare ne rende più facile e meno traumatico il passaggio attraverso
l'istmo delle fauci e l'esofago.
Regolazione della secrezione salivare
La secrezione salivare é esclusivamente sotto controllo nervoso. Le ghiandole
sono innervate sia da nervi colinergici parasimpatici che da nervi adrenergici
ortosimpatici. Inoltre, tecniche immunoistochimiche hanno dimostrato la
presenza, nelle terminazioni nervose, di VIP e di sostanza P. Le fibre nervose
colinergiche postgangliari, parasimpatiche, si portano alle cellule sia delle
estremità secretorie che dei dotti e la loro stimolazione induce la più
elevata produzione di secrezione salivare. La stimolazione simpatica, invece,
causa un incremento della secrezione salivare di modesta e variabile entità.
Anche le cellule mioepiteliali sono innervate dal simpatico e la risposta
secretoria può, almeno in parte, essere mediata dal loro contributo.
La secrezione salivare é totalmente riflessa ed involontaria (si può aumentare
volontariamente la secrezione solo sfruttandone i meccanismi riflessi). Le
strutture nervose che presiedono a tale regolazione sono situate a livello
bulbare. Si distinguono nuclei salivatori superiori (prevalentemente interessati
alla parotide) ed inferiori (prevalentemente interessati alla sottomandibolare
ed alla sottolinguale).
Come già riferito, la regolazione della secrezione dipende interamente dal
sistema nervoso parasimpatico e da quello ortosimpatico. Il sistema nervoso
ortosimpatico ha estensione toraco-lombare e quindi arriva alle ghiandole
salivari tramite i nervi toracici più alti, con fibre che non si interrompono
subito a livello gangliare ma si portano in alto, fino al ganglio cervicale
superiore dove, interrompendosi, prendono connessione sinaptica con fibre
postgangliari che, a loro volta, si portano alle tre ghiandole. La innervazione
parasimpatica esce dai gangli del V, VII e IX nervo cranico e si porta a tutte
le ghiandole subito o tramite i gangli otico o mascellare, molto vicini agli
organi innervati.
Esistono tre fasi nella secrezione salivare:
cefalica, orale e gastrica.
La fase cefalica della secrezione salivare
é stata ampiamente documentata da Pavlov con i suoi esperimenti sui riflessi
condizionati. Pavlov induceva salivazione in animali da laboratorio associando
alla somministrazione di cibo stimoli (acustici o visivi) che di per sé non
avevano alcuna correlazione di tipo alimentare (suoni di campanelli, accensione
o spegnimento di lampadine) dimostrando che esiste, in assoluta mancanza di
stimolazioni orali, un controllo corticale, o comunque certamente sopra-bulbare,
della funzione salivatoria. Stimoli certamene non salivatori possono quindi
essere in grado di promuovere la secrezione salivare se associati, per un
sufficiente periodo di tempo, a stimoli direttamente salivatori, quali la
presenza del cibo nella cavità orale. Una volta instauratasi, la salivazione
cefalica dovuta ad un certo stimolo, se non sostenuta da una successiva
assunzione di cibo, tende lentamente ad estinguersi.
Dopo la fase cefalica, con l'assunzione del cibo,
si manifesta la fase orale, dovuta alla
stimolazione meccanica dei recettori tattili presenti nella mucosa della cavità
orale che, in via riflessa breve, incrementa la secrezione salivare. Sono più
efficaci, stranamente, i corpi lisci mentre lo sono meno le sostanze a
superficie ruvida. Un altro stimolo che a livello della cavità orale induce
abbondante salivazione è l'acidità del cibo introdotto. Tale riflesso,
palesemente protettivo, determina diluizione del contenuto orale, diminuendone
il potere nocivo nei confronti della mucosa, anche esofagea. Sostanze in grado
di evocare precise sensazioni gustative (salato, amaro o dolce) non presentano
una rilevante influenza sull'attività salivatoria.
Nell'ultima fase, la fase
gastrica, la presenza di contenuto nello stomaco stimola, per via
nervosa riflessa, il mantenimento della secrezione salivare che così
contribuisce alla ulteriore diluizione del pasto.
Nella fase cefalica, la salivazione é apparentemente interamente sotto il
controllo parasimpatico. Nella fase orale, essa é mediata da riflessi locali
verosimilmente ancora parasimpatici ed infine, nella fase gastrica, alla
componente parasimpatica si associa l'azione di una notevole componente
ortosimpatica (aumento del flusso ematico alle ghiandole salivari) che, in
condizioni normali, promuove anch'essa l'attività salivatoria, contrariamente a
quanto si verifica in altri distretti, dove l'azione parasimpatica e quella
ortosimpatica sortiscono effetti opposti. Tuttavia si possono avere azioni
"paradosse" del sistema ortosimpatico che si manifestano con
vasodilatazioni abnormi a livello ghiandolare, in assenza di contemporanea
stimolazione parasimpatica, determinando una anomala secchezza delle fauci. Ad
ogni modo, qualunque stimolazione intensa del sistema nervoso centrale (paura,
emozioni, stress) è in grado di produrre in via riflessa, attraverso le
terminazioni nervose del sistema neurovegetativo o mediatori chimici o ormonali
riversati nel torrente circolatorio, variazioni dell'attività delle ghiandole
salivari. Capita anche che, in situazioni di stress, la ventilazione diventi
spesso orale (anzicché nasale) ed aumenti di intensità, determinando maggiore
evaporazione di liquidi dalla cavità orale. Non è quindi certo che in tutti i
casi la responsabilità della secchezza delle fauci sia da attribuirsi
esclusivamente al blocco o alla inibizione della funzione secretoria salivare.
Fisiopatologia della secrezione salivare
La secrezione salivare può essere temporaneamente inibita da infezioni o da
farmaci quali gli agenti anticolinergici. Inibizioni più durature si verificano
in seguito ad irradiazioni della testa o del collo per la cura di tumori. Danni
permanenti si hanno nella sindrome di Sjögren, un'affezione mediata dal sistema
immunitario. In questi casi si hanno secchezza della bocca (xerostomia),
difficoltà del linguaggio e della deglutizione, vaste carie dentali e disturbi
del gusto.
MASTICAZIONE
(Prima parte)
La masticazione è un complesso, armonico e delicato meccanismo che, per
compiersi efficacemente, necessita del contributo di fattori ossei, fattori
articolari, fattori occlusali, fattori muscolari e fattori nervosi.
Fattori ossei
Dotto
ressa
Fra le componenti ossee dell'apparato stomatognatico, la mandibola è
quella che riveste il maggiore interesse dal punto di vista funzionale,
costituendone il principale osso mobile. Per le altre ossa (mascellari,
temporali, palatine ed osso ioide) si rimanda a fonti più appropriate.
La mandibola è un osso impari e mediano, la cui sagoma ad arco parabolico
richiama l'immagine di un ferro di cavallo. La si può immaginare anche come un
ponte osseo con le spalle nelle articolazioni temporomandibolari e la chiave di
volta del suo arco nella sinfisi mentoniera.
Alla nascita, la mandibola è ancora suddivisa in
due parti da una fessura mentoniera. Le due parti si uniscono successivamente e
della fusione che ne deriva rimangono tracce visibili in una o più spine
mentali, piccole apofisi poste sulla faccia posteriore della mandibola: sono
spesso visibili nelle radiografie della mandibola scattate in proiezione
assiale. Nella faccia esterna, o anteriore, del mento si trovano costantemente
due tubercoli mentonieri, visibili anch'essi nelle radiografie assiali, ai due
lati del mento. Alcuni ossicini mentonieri possono persistere nell'adulto. Dalla
estremità posteriore del corpo della mandibola, la porzione orizzontale
dell'osso, emergono due rami montanti, pari e simmetrici, diretti verso l'alto e
posteriormente con andamento speculare, formando con il corpo i due angoli
mandibolari, ottusi. Alla nascita, l'angolo mandibolare misura circa 150° e si
riduce nell'adulto a circa 130°. Le variazioni minime e massime nell'ambito
fisiologico sono, per l'adulto, di 90° e 140°. Ogni ramo montante della
mandibola termina in alto e posteriormente con un processo condiloideo, la cui
testa si articola con l'osso temporale nell'articolazione temporomandibolare. Il
processo condiloideo è separato in avanti dal processo coronoideo, sede di
impianti muscolari, tramite la incisura sigmoidea. Sulla faccia esterna del ramo
montante della mandibola si trova una tuberosità masseterina per l'impianto del
muscolo massetere; sulla faccia interna è presente una tuberosità pterigoidea
e poi, sul corpo della mandibola, una fossa digastrica per il muscolo omonimo.
Sopra la fossa digastrica corre la linea miloioidea, per il muscolo omonimo, che
segnala al radiologo il confine caudale della cavità orale: il muscolo
miloioideo, infatti, forma il pavimento della cavità orale.
I denti sono costituiti fondamentalmente
dalla dentina. La porzione scoperta del dente, corona, è rivestita dallo
smalto; la porzione alveolare, radice, è ricoperta dal cemento. La zona di
giunzione fra smalto e cemento, colletto, è ricoperta dalla mucosa gengivale.
La cavità del dente, occupata dalla polpa dentaria, comunica con l'esterno
attraverso il foro radicolare, in cui passano i vasi sanguigni e le fibre
nervose. La dentina è simile all'osso, ma più resistente. E' attraversata dai
tubuli dentinali, che si portano radialmente dalla superficie pulpare, ove hanno
un diametro di 3 - 4 micron, alla giunzione con lo smalto (giunzione
amelo-dentinale) ove il loro diametro si riduce a circa 1 micron. Dai tubuli
partono branche laterali (canalicoli), del diametro di 1 micron, che formano
anastomosi con altri tubuli. La dentina è pertanto permeata da una rete di
tubuli e canalicoli. Nei tubuli sono contenuti sia liquido extracellulare che i
processi citoplasmatici delle cellule che provvedono a formare ed a mantenere la
dentina stessa (odontoblasti). I corpi cellulari degli odontoblasti sono
localizzati nella polpa e formano uno strato che riveste la superficie pulpare
della dentina. Alcuni processi degli odontoblasti, specie nel dente giovane, si
dividono, in prossimità della giunzione amelo-dentinale, in branche terminali,
alcune delle quali possono penetrare nei fusi dello smalto. Non tutti i processi
degli odontoblasti giungono fino alla parte terminale dei tubuli.
Lo smalto è costituito per 96-98% da materiale inorganico ed è assolutamente
privo di cellule e di terminazioni nervose. Negli strati più interni, a livello
della giunzione amelo-dentinale, si osservano prolungamenti dei tubuli dentinali
chiamati, per la loro forma, fusi dello smalto, conteneti la parte terminale dei
processi odontoblastici. Lo smalto ha una certa permeabilità per sostanze
organiche di basso peso molecolare, comportandosi dal punto di vista fisico come
una membrana semipermeabile.
Il cemento, simile all'osso compatto, riveste la radice del dente e costituisce
il tramite per il quale il dente viene unito all'osso alveolare dal legamento
periodontale. Il cemento è avascolare e privo di innervazione.
La polpa si divide in una porzione parietale (zona odontogenetica) ed in una
porzione centrale. La prima è costituita dallo strato delle cellule
odontoblastiche che tappezzano la cavità pulpare, da una zona intermedia priva
di cellule e da una zona più interna, ricca di cellule fibroblastiche e
mesenchimali. La porzione centrale contiene fibre nervose e vasi sanguigni
immersi nel connettivo.
Fattori articolari
Per la forma a ferro di cavallo della mandibola, le
articolazioni
temporomandibolari (ATM) sono uniche nel corpo umano. Compongono
infatti una doppia giuntura articolata posta tra i due condili dell'osso
mandibolare - quello di destra e quello di sinistra - e due aree circoscritte
delle ossa temporali. Queste ultime corrispondono ai tubercoli articolari ed
alle due fossette del temporale, dette mandibolari. L'insieme funzionale delle
due ATM viene dunque globalmente a costituire una articolazione diartroidale
bicondilea nella quale due settori anatomicamente separati - quello di destra e
quello di sinistra - sono anatomicamente e funzionalmente uniti e sono resi
sincronicamente interdipendenti. Ogni movimento della mandibola coinvolge
necessariamente entrambe le articolazioni temporomandibolari, anche se non
obbligatoriamente allo stesso modo.
Le due teste condiloidee della mandibola, congiunte dal collo alla branca
montante, sono poste all'apice dell'angolo postero-superiore di tali apofisi.
Una faccetta articolare copre l'intera faccia superiore della testa del condilo,
un breve tratto del versante anteriore ed un tratto del versante posteriore
lungo circa mezzo centimetro. La faccetta articolare è rivestita da uno strato
fibrocartilagineo povero di condrociti: tra le articolazioni diartroidali, le
ATM sono uniche nell'avere faccette articolari rivestite non da cartilagine
jalina, ma da fibrocartilagine.
Nella fossa mandibolare dell'osso temporale è predisposta una faccetta
articolare ovale, decisamente concava, per accogliere un disco fibrocartilagineo
e la testa del condilo mandibolare, delimitata in avanti dal tubercolo
articolare del processo zigomatico. Anche la faccetta articolare temporale è
rivestita da uno strato di tessuto fibrocartilagineo. Nella porzione più
posteriore, non articolare, della fossa mandibolare dell'osso temporale - tra
faccetta articolare e forame auricolare - trova sovente posto tessuto epiteliale
ghiandolare della ghiandola salivare
parotidea.
Il processo zigomatico dell'osso temporale è una sottile e lunga prominenza
ossea che assume, nell'insieme, la sagoma di un prolungato emicilindro,
sporgente in avanti e diretto quasi orizzontalmente. La faccia inferiore
dell'arco zigomatico forma la porzione anteriore dell'area articolare della
temporomandibolare: pertanto tale area è rivestita anch'essa da tessuto
fibrocartilagineo.
Il tubercolo articolare si identifica con la radice anteriore
della faccia inferiore del processo zigomatico dell'osso temporale. La
prominenza determinata dall'osso zigomatico costituisce un importante punto di
riferimento nella semeiotica del volto. Il tubercolo dello zigomatico è infatti
posto dove la radice anteriore si incontra con la radice posteriore della faccia
inferiore dell'apofisi. Sul tubercolo si inserisce il capo
prossimale del
legamento temporomandibolare.
Un disco fibrocartilagineo (menisco articolare) divide la cavità articolare
della ATM in due compartimenti separati: uno superiore ed uno inferiore. Il
primo si trova tra la faccetta articolare dell'osso temporale ed il disco; il
secondo tra la faccia inferiore del disco ed il condilo della mandibola. Il
disco fibrocartilagineo ha un perimetro grossolanamente ovale ed una faccia
inferiore concava, che si adatta alla superficie convessa del condilo
mandibolare. La faccia superiore del disco è invece concavo-convessa, sellare
(dall'avanti all'indietro) ed è morfologicamente definibile come l'immagine
negativa della superficie della porzione anteriore della fossa mandibolare e del
tubercolo articolare del temporale.
Fibre del disco articolare si inseriscono, in avanti, sul capo superiore del
muscolo pterigoideo esterno (laterale) atraverso la parete anteriore della
capsula; il capo inferiore dello pterigoideo laterale collega invece la
superficie esterna del processo pterigoideo dello sfenoide alla fossa
pterigoidea del collo del condilo. (Questi due capi funzionano in realtà come
muscoli distinti ed antagonisti: il capo inferiore si contrae durante l'apertura
della bocca, il capo superiore durante la chiusura). Il restante del contorno
meniscale aderisce fortemente alla capsula fibrosa dell'articolazione, più
saldamente in direzione nucale. Il disco è anche agganciato, con robusti tralci
fibrosi, ai margini mediale e laterale del condilo mandibolare. Tali legami
creano un'unione solidale tra disco fibrocartilagineo e condilo della mandibola,
perciò le due formazioni anatomiche si spostano insieme nei movimenti di
protrusione e di retrusione della mandibola. Lo spessore del disco
fibrocartilagineo è minore al centro, nella cosiddetta zona intermedia; è
massimo in altre due zone: la banda anteriore e la banda posteriore. Davanti
alla banda anterore c'è il prolungamanto anteriore, che raggiunge, come detto,
il muscolo pterigoideo esterno: si collega, a mezzo di rami fibrosi, sia con la
mandibola, sia col temporale. La regione bilaminare è situata dietro la banda
posteriore del disco ed è chiamata così perchè è dissociabile in due strati
laminari, uno superiore ed uno inferiore, separati da un plesso venoso. Le fibre
dello strato laminare superiore raggiungono l'osso temporale, quelle dello
strato inferiore si inseriscono sull'osso mandibolare. Per la sua complessa
morfologia, il disco fibrocartilagineo della ATM assume, nel suo insieme, un
particolare aspetto: è stato paragonato ad un berretto con visiera o ad un berretto da fantino.
L'involucro fibroso della capsula articolare segue, con la sua porzione distale,
il contorno del collo del condilo. Sull'osso temporale, la linea prossimale di
inserimento circonda la fossa glenoidea, correndo lungo il bordo del tubercolo
anteriore, sul contorno della fossa mandibolare ed in prossimità della fessura
timpanico-squamosa. Il contenimento contro il temporale dell'osso mandibolare è
dovuto alla capsula fibrosa, rinforzata da tre importanti ligamenti: il
legamento temporomandibolare, che corre sotto la ghiandola parotide e si
inserisce in alto sul processo zigomatico ed il basso sul collo del condilo
mandibolare, esternamente e posteriormente; il legamento sfenomandibolare,
alloggiato in un piano più profondo, lungo la faccia interna della capsula
fibrosa: origina dalla spina dello sfenoide e raggiunge in basso la lingula del
foro mandibolare; il legamento stilomandibolare, che ha un
posizionerelativamente lontana dalla capsula fibrosa: origina in alto dall'apice
del processo stiloiodeo e raggiunge, in basso, il margine posteriore del ramo
montante della mandibola, in vicinanza dell'angolo. La capsula è piuttosto tesa
nella sua porzione inferiore, dove il disco fibrocartilagineo e la testa del
condilo sono collegati dalle connessioni fibrose su menzionate; è invece lassa
al disopra del disco. Ciò consente quella particolare mobilità
dell'articolazione che porta all'apertura ed alla chiusura della bocca. Nel
corso del primo dei due movimenti, infatti, il collo mandibolare ed il disco,
mentre ruotano, scivolano in avanti; nella chiusura della bocca si riportano
entrambi nella posizione di partenza, scivolando sulla superficie
fibrocartilaginea della faccetta articolare del temporale.
Come in ogni altra articolazione diartroidale sana, due o tre strati di cellule
sinoviali rivestono la parete interna della capsula fibrosa, costituendo una
membrana sinoviale. Rivestono anche parte del condilo mandibolare e parte del
tendine del muscolo pterigoideo esterno. La membrana sinoviale si riflette,
inoltre, sulla faccia superiore e sulla faccia inferiore del disco
fibrocartilagineo. Perciò in ogni articolazione temporomandibolare esistono due
cavità sinoviali distinte: un compartimento sinoviale superiore, o
temporo-discale, ed un compartimento sinoviale inferiore, o disco-mandibolare.
Sotto il profilo funzionale, le due componenti parietali del compartimento
superiore - cioè la fossetta articolare del temporale ed il disco
fibrocartilagineo - si muovono reciprocamente per scivolamento dell'una
sull'altra. Il compartimento inferiore si raffigura funzionalmente, invece, come
un'articolazione a cardine, cioè come un ginglimo.
L'ATM riceve sangue da rami della arteria temporale superiore e dalla
mascellare. E' innervata da ramificazioni della branca mandibolare del
trigemino.
Il legamento periodontale o paradonzio
comprende un insieme di tessuti connettivali che consentono il fissaggio dei
denti alle ossa mascellari.
Anatomicamente, il fissaggio del dente all'alveolo è considerato
un'articolazione chiamata gonfosi, caratterizzata da minimo movimento delle
parti. Il legamento periodontale occupa lo spazio compreso tra il cemento e la
superficie dell'osso alveolare e si estende, verso la corona, fino alla lamina
propria della gengiva. Le fibre collagene del legamento si inseriscono da un
lato nel cemento e dall'altro nel periostio dell'osso alveolare. Costituito da
tessuto molle, funziona quindi come mezzo di unione tra due tessuti
mineralizzati, garantendo contemporaneamente al dente sia la fissità
nell'alveolo che un certo grado di spostamento in seguito alle sollecitazioni
verticali ed orizzontali. Le sollecitazioni orizzontali a direzione mesiale e
distale provocano movimenti pressocchè nulli del dente, se questo è a contatto
coi denti contigui; quelle orizzontali dirette in senso vestibolare o linguale
fanno sì che il dente agisca sull'alveolo come una leva, generando zone di
pressione e di trazione.
Modellamento e rimodellamento dei capi articolari.
Il condilo mandibolare è formato da osso spugnoso ricoperto in superficie da
una corteccia di osso compatto, di spessore variabile, a sua volta rivestita da
cartilagine fibrosa. La porzione temporale dell'articolazione è strutturata in
modo analogo. La cavità glenoidea presenta nella sua parte più profonda solo
una teca di osso compatto relativamente sottile. Sia le strutture calcificate
che quelle molli possono andare incontro a notevoli cambiamenti
morfo-strutturali per rimodellamento. Questo processo segue immediatamente il
modellamento che si è avuto durante l'accrescimento.
Il processo di accrescimento non è solo espansivo, ma conduce ad un cambiamento
di forma delle strutture interessate, con un meccanismo di addizione
differenziata e di rimozione di materiale in determinate zone. Quando nuovo osso
si sviluppa in un segmento osseo, la posizione relativa dei segmenti limitrofi
viene alterata. La deposizione di nuovo tessuto osseo modifica la forma del
segmento interessato per adattarla alla nuova posizione e accrescerne le
dimensioni. Il processo di accrescimento è direttamente influenzato, oltre che
da fattori endocrini, dalle forze che si applicano sull'osso.
|
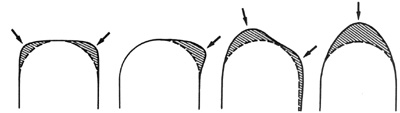
|
| Disegno schematico delle diverse
forme assunte dalle sezioni condilari a seconda della distribuzione del
rimodellamento (zone tratteggiate). Le frecce indicano i punti di più
intensa attività rimodellante. |
Se ad esempio nel corso
dell'accrescimento il muscolo temporale subisce una lesione, il processo
coronoideo della mandibola non si sviluppa; in caso di edentulismo congenito,
invece, non si sviluppa il processo alveolare. Le forze pressorie possono
condurre ad accrescimento della cartilagine, mentre provocano riassorbimento del
tessuto osseo, che si accresce invece in presenza di forze di trazione.
La componente ossea dell'ATM è sede di attivo rimodellamento dovuto alla
progressiva sostituzione dell'osso primario con un sistema di osteoni secondari
di successive generazioni e di brecce ossee concentriche. Se il processo rimane
interno al profilo originario muta le caratteristiche biomeccaniche del segmento
osseo interessato, mentre se si sviluppa verso l'esterno conduce anche ad un
cambiamento della forma. In tutti i casi, il rimodellamento rappresenta un
adeguamento alle necessità funzionali conseguente alle forze applicate sull'osso.
Sembra che, se sottoposto a stress, l'osso generi cariche elettriche, sia in
vivo che in vitro. L'origine di tali cariche, però, è ancora in discussione:
non è chiaro se sia imputabile ad attività piezoelettrica del collagene e/o
dei cristalli sottoposti a stress o se sia il prodotto della normale attività
metabolica. Forze pressorie esercitate alle estremità di un osso lungo
provocano l'insorgenza di una carica elettrica negativa sulla parte che tende a
concavizzarsi e di una carica positiva sul lato convesso. Ciò indurrebbe un
rimodellamento dell'osso verso il lato concavo (che però non è quello
sottoposto a trazione), tendente a neutralizzare le forze applicate.
Il continuo processo di assorbimento e deposizione, che caratterizza il
rimodellamento, mantiene in stato dinamico l'osso dal termine dell'accrescimento
fino alla morte. A seconda del prevalere dell'assorbimento o della deposizione,
si parla di rimodellamenro "progressivo" o "regressivo".
Nell'ATM il rimodellamento osseo si ritrova in tutte le età dell'individuo
adulto. Nel condilo mandibolare, dopo una intensa "progressione" tra i
18 e 25 anni, gli indici di rimodellamento rimangono più o meno costanti in
tutte le età successive. La distribuzione del rimodellamento condilare non è
casuale, bensì mirata con prevalenza in determinate zone: le parti anteriori e
posteriori del condilo sono più frequentemente interessate, la parte superiore
più raramente. La diversa distribuzione del rimodellamento determina una
notevole varietà morfologica tra un condilo e l'altro e tra le diverse sezioni
dello stesso condilo. Si osserva altresì una stretta relazione morfo-funzionale
tra le strutture di primo ordine (trabecole e osso compatto) visibili
macroscopicamente e quelle di secondo ordine (lamelle ed osteoni) visibili al
microscopio: le trabecole tendono ad allinearsi perpendicolarmente alle zone di
più intenso rimodellamento. In alcuni casi le lamelle trabecolari attraversano
l'osso compatto fino in prossimità della superficie esterna; altre volte si
interrompono in corrispondenza della zona di rimodellamento o si sventagliano
determinando l'aspetto di un supporto a calice aperto verso questa zona.
Modellamento e rimodellamento dei tessuti molli
articolari.
Il tessuto non calcificato che riveste il condilo e la parte temporale dell'ATM
è costituito da diversi strati: strato fibroso verso la superficie articolare,
strato proliferativo (o mesenchima indifferenziato), strato intermedio (poco
distinto) e strato cartilagineo. Il suo spessore varia nelle diverse zone ed è
maggiore nelle parti anteriori e antero-superiori del condilo e nella parte
posteriore ed inferiore del tubercolo articolare. Di norma ha un aspetto liscio
ma, osservato al miscroscopio a scansione, presenta spesso piccole depressioni o
fossette che gli conferiscono un aspetto di palla da golf; queste depressioni
sono di più frequente riscontro nella zona media antero-superiore della
superficie condilare. Cambiamenti di forma, visibili macroscopicamente, dovuti a
localizzati cambiamenti di spessore dei tessuti molli articolari, sono di
osservazione relativamente frequente nelle ispezioni anatomo-patologiche. Alla
base di questi cambiamenti di forma vi sarebbe una stimolazione del mesenchima
indifferenziato e la sua conseguente differenziazione in cartilagine.
I cambiamenti poco marcati della superficie articolare, non visibili ad occhio
nudo ma osservabili al microscopio a scansione, possono non interessare il
sottostante tessuto osseo, mentre le depressioni della cartilagine visibili
macroscopicamente sono accompagnate da analogne depressioni del tessuto osseo.
Tali riassorbimenti ossei localizzati rappresentano una forma di rimodellamento
regressivo e sono particolarmente frequenti sulla superficie condilare
posteriore della mandibola.
Fattori occlusali
Col termine di occlusione o combaciamento dentario si intende il modo in cui le
arcate dentarie vengono tra di loro in contatto ed in quale modo ed in quale
posizione dello spazio rimangono in contatto. Al fenomeno occlusale partecipano
l'apparato dento-parodontale, l'articolazione temporomandibolare ed il sistema
neuro-muscolare. Il primo rappresenta la parte più interessata al combaciamento
e quindi all'occlusione; l'articolazione è il collegamento fra la parte mobile
(mandibola) e la parte fissa (cranio); il sistema neuromuscolare costituisce la
parte dinamica, quella cioè veramente attiva, che permette il movimento.
Da un punto di vista pratico, l'occlusione ideale si ha quando in due arcate in
occlusione:
1) la superiore sia più grande dell'inferiore di una mezza cuspide nel senso
della larghezza (diametro trasverso);
2) i denti frontali superiori avanzino e coprano gli inferiori per un terzo
circa della loro altezza, nel senso della lunghezza (diametro antero-posteriore
o sagittale);
3) i primi molari permanenti, che rappresentano la chiave dell'occlusione, siano
in rapporto tale che, sul piano sagittate, gli inferiori avanzino di mezza
cuspide rispetto ai superiori.
In tal modo, ogni dente di una arcata articola con due denti dell'arcata
antagonista, ad eccezione degli ultimi molari superiori e dei due incisivi
centrali inferiori che articolano con un solo antagonista.
Le arcate dentarie vengono normalmente in contatto in posizioni diverse. In
posizione di massima intercuspidazione (ICP) si ha il massimo avvicinamento
spaziale tra mandibola e mascella con il massimo numero di contatti
interdentari, posizione che viene spesso raggiunta alla fine del ciclo
masticatorio e durante la deglutizione. Nel caso di arcate normali ed integre,
le cuspidi palatine superiori e quelle vestibolari inferiori contattano in ICP
con una fossa antagonista o con creste marginali di denti contigui: cuspidi
"di stampo" o "di appoggio". Una occlusione in cui tutte le
cuspidi di appoggio contattano con una fossa di un dente antagonista senza
impegnarsi negli spazi interdentari viene detta occlusione organica.
Il contatto interdentale avviene in circa l'80% dei cicli masticatori ma,
ovviamente, la frequenza dipende dalla natura del cibo che viene masticato. Nei
primi cicli di una sequenza è più raro, ma nelle fasi terminali, quando la
consistenza fisica del cibo è stata sufficientemente modificata, il contatto
avviene costantemente alla fine di ciascun ciclo. Tale contatto, tuttavia, non
é statico, non avviene cioè quando la mandibola è ferma. Nella fase finale di
un ciclo masticatorio, infatti, le superfici occlusali dei denti inferiori si
muovono in contatto con le superfici occlusali dei denti superiori (contatto
mobile), lungo una via determinata dalla forma delle cuspidi dentali. Quando i
denti sono in ICP, il contatto da mobile diventa statico. In genere, il contatto
mobile durante la fase di lavoro ha un'escursione di 1 mm circa, ma l'ampiezza
dipende dal tipo di movimento (verticale o rotatorio): quando è richiesa
un'azione di macina può arrivare a 2 mm. Un certo grado di contatto mobile tra
le superfici occlusali avviene anche all'inizio della fase di apertura, quando
il movimento inizia con una deviazione contralaterale al lato di lavoro.
In fase dinamica di crescita i contatti interdentari svolgono un ruolo
morfogenetico di primaria importanza. Il combaciamento interdentario, infatti,
comporta l'insorgenza di stimoli che, tramite il sistema propriocettivo, si
portano a livello mesencefalico, potendo da qui influenzare l'attività
muscolare masticatoria.
Uno schema di contrazione armonica dei muscoli masticatori si trova associato ad
un rapporto occlusale ottimale, in cui idealmente la posizione dei condili è
nel centro della fossa temporale. La interdigitazione delle cuspidi dentali
determinerà quindi la posizione della mandibola quando le arcate vengono a
contatto, e la posizione dei condili in relazione alle fosse dipenderà dalla
intercuspidazione. Gli schemi di chiusura vengono memorizzati dal sistema
nervoso, purché i segnali generati dalle posizioni di contatto vengano ripetuti
di frequente, come accade nella fase della deglutizione. Nel soggetto sano la
posizione di intercuspidazione massima è molto stabile, pertanto i movimenti di
chiusura portano ad un rapporto di occlusione centrica. Ciò comporta che la
posizione muscolare di contatto è raggiunta con il minimo sforzo muscolare. Se
vi è un'alterazione della intercuspidazione massima, in quanto la postura della
mandibola è alterata da precontatti o da squilibri occlusali, per raggiungere
la posizione muscolare di contatto vengono richiesti sforzi muscolari
addizionali e la posizione occlusale risulterà stabilizzata dai soli muscoli,
ai quali vengono così richieste tensioni eccessive.
Movimenti
della mandibola.
MASTICAZIONE
(Seconda parte)
Movimenti della
mandibola.
I movimenti della mandibola sono tutti bicondilari, in quanto vengono sempre
impegnate contemporaneamente le articolazioni dei due lati. Possono avvenire per
rotazione e/o translazione condilare, nei tre piani dello spazio: sagittale
mediano, frontale ed orizzontale, fra loro ortogonali. Per seguire i movimenti
elementari che la mandibola compie nello spazio occorre quindi definire i
singoli piani dello spazio a cui si fa riferimento e gli assi intorno ai quali
avvengono i movimenti di rotazione dei condili. Il piano sagittale, disposto in
senso verticale ed antero-posteriore, in realtà non è unico: sono infatti
infiniti i piani sagittali che attraversano il cranio con direzione verticale ed
andamento antero-posteriore. Il piano sagittale mediano è invece ben definito,
in quanto è l'unico che, passando per la sutura intraparietale, divide il
cranio in due metà speculari, destra e sinistra. Anche i piani frontali ed i
piani orizzontali sono infiniti per definizione e dividono il cranio,
rispettivamente, in piani anteriori e posteriori o superiori ed inferiori. Per
quanto riguarda gli assi di rotazione dei condili, i movimenti articolari
possono avvenire intorno a tre assi: orizzontale, verticale ed antero-posteriore
di cui uno solo, quello orizzontale, è comune ad uno dei due condili: è
bicondilare. Quando i condili ruotano intorno all'asse bicondilare, il loro
movimento è sempre consensuale e la mandibola si muove nel piano sagittale. Se
la rotazione avviene intorno ad uno degli assi verticali od anteroposteriori si
ha invece rotazione di un solo condilo alla volta (condilo ruotante), mentre il
condilo contralaterale esegue un movimento translatorio od orbitante descrivendo
un piccolo arco di cerchio (condilo orbitante). La rotazione dei condili attorno
agli assi verticali provoca movimenti sul piano orizzontale, quella attorno agli
assi antero-posteriori provoca movimenti sul piano frontale.
|
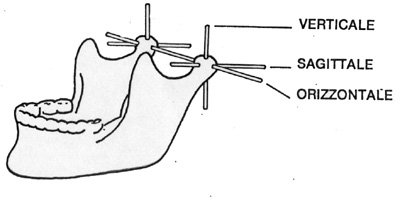
|
| Posizione dei cinque principali
assi di rotazione della mandibola. |
I movimenti elementari della mandibola
sono di tre tipi, di seguito descritti:
1) abbassamento ed innalzamento, chiamati anche, rispettivamente, di apertura e
chiusura;
2) proiezione in avanti e di retrazione in dietro, chiamati anche,
rispettivamente, di protrusione e retrusione;
3) lateralità.
Movimenti di apertura e di chiusura.
I movimenti di abbassamento ed innalzamento della mandibola sono movimenti
semplici, che vengono eseguiti nel piano sagittale. Entrambi si attuano per sola
rotazione dei condili attorno all'asse bicondilare o per movimenti combinati di
rotazione e di traslazione dei condili. Il movimento di abbassamento può
iniziare dalla occlusione centrica (massimo contatto - intercuspidazione - delle
superfici occlusali dei denti delle arcate antagoniste) o dalla posizione di
riposo (leggero grado di apertura della mandibola, per la presenza di un piccolo
spazio tra le superfici occlusali delle arcate contrapposte). Il passaggio dalla
occlusione centrica alla posizione di riposo avviene per rotazione di 2° circa
della testa del condilo attorno all'asse bicondilare, con impegno della sola
articolazione condilo-discale. Partendo da una delle due posizioni, il movimento
di apertura della mandibola può essere scomposto in due fasi fondamentali: di
rotazione e di translazione dei condili. Il movimento di rotazione attorno
all'asse bicondilare è chiamato anche "a cerniera" e l'asse
bicondilare "asse cerniera". Nel corso di questo movimento, i condili
ruotano di circa 10° - 12° in senso antiorario consentendo una distanza di 2 -
2,5 cm tra i margini degli incisivi superiori ed inferiori. Nel corso della
rotazione, il movimento traslatorio dei condili è minimo (1 - 2 mm). In questa
fase i condili possono mantenere la relazione centrica (posizione più retrusa
non forzata). L'ulteriore abbassamento della mandibola avviene per traslazione
bilaterale dei condili che si spostano consensualmente in basso ed in avanti,
lungo la cosiddetta via o tragitto condilare, fino sul tubercolo articolare del
temporale. Nel movimento di traslazione sono impegnate entrambe le articolazioni
del disco articolare, che viene spostato in avanti anche per contrazione del
muscolo pterigoideo esterno. Raggiunta questa posizione, la testa del condilo
può eseguire un ulteriore movimento rotatorio che lo sposta fin quasi all'apice
del tubercolo articolare. Nella massima apertura, lo spazio tra gli incisivi
superiori ed inferiori è di 5 - 6 cm.
Nel movimento di innalzamento della mandibola, che inizi dalla posizione di
massima apertura, avviene dapprima un movimento semplice di rotazione del
condilo in senso inverso, mentre il disco articolare è ancora trattenuto in
avanti e, infine, un movimento combinato di rotazione e traslazione del condilo
che si sposta posteriormente, con contemporaneo scivolamento del disco
articolare che rientra nella cavità del temporale.
Movimenti di protrusione e retrusione.
Sono movimenti che avvengono nel piano orizzontale, con scivolamento dell'arcata
dentaria inferiore sotto quella superiore. Il movimento si attua per semplice
traslazione dei condili, con minima rotazione (inferiore a 1°). Nella
protrusione, la testa del condilo ed il disco articolare scorrono sulla
superficie del tubercolo articolare del temporale portandosi in avanti ed in
basso, fino all'apice del tubercolo stesso. I movimenti condilari sono
bilaterali e contemporanei. La traslazione dei condili nella protrusione ha
circa la stessa ampiezza che ha nell'apertura massima della mandibola. Nella
protrusione estrema, gli incisivi inferiori oltrepassano in avanti quelli
superiori di alcuni mm (7 - 10). Nella retrusione avviene il movimento inverso.
Movimenti di lateralità. Sono movimenti che
avvengono sul piano orizzontale e consentono lo spostamento della mandibola
verso destra e verso sinistra. Possono avvenire a mandibola chiusa, con
scorrimento delle superfici occlusali dei denti contrapposti, oppure a mandibola
aperta, ma solo nella fase di apertura a cerniera, quando i condili sono
provvisti di mobilità nelle cavità articolari. Nella fase di massima apertura,
quando i condili sono contrapposti al tubercolo temporale, i movimenti laterali
sono minimi. Nei movimenti di lateralità i condili dei due lati si comportano
in modo diverso: un condilo ruota attorno all'asse verticale (condilo ruotante)
mentre il controlaterale subisce un movimento di traslazione descrivendo un
piccolo arco di cerchio (condilo orbitante). Ad esempio, nello spostamento della
mandibola verso destra, il movimento avviene per rotazione del condilo destro,
che funge da perno, attorno ad un asse verticale che in realtà è situato
posteriormente al condilo. Il condilo ruotante subisce sempre una leggera
rotazione con spostamento laterale e posteriore. Il condilo orbitante viene
invece spostato in avanti e medialmente e si pone sotto il tubercolo articolare
del temporale. Il movimento laterale della mandibola è anche chiamato
abduzione, il ritorno alla posizione di riposo adduzione.
Nella escursione laterale della mandibola si verifica uno scorrimento delle
cuspidi di appoggio lungo i versanti di cuspidi antagoniste (cuspidi "di
taglio" o "di guida"). Nel caso che durante l'escursione in
lateralità contattino solo i canini si parla di "guida canina",
altrimenti di "contatto di gruppo". Dal lato opposto al movimento può
non esserci alcun contatto dentario o può esserci contatto fra una o più
coppie di denti antagonisti ("bilanciamento").
Traiettorie.
La combinazione dei movimenti verticali di apertura e di chiusura con i
movimenti alternati verso destra e verso sinistra dà luogo a movimenti di
rotazione della mandibola. Questi, che sono quindi la risultante dei movimenti
elementari eseguiti sul piano sagittale ed orizzontale, descrivono una
traiettoria, circolare od ellittica, sul piano frontale, suddivisibile in sei
componenti sul piano sagittale.
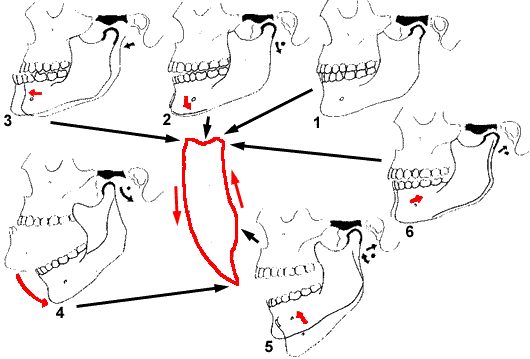 |
| Traiettoria della mandibola, sul
piano sagittale mediano. |
Le metodiche più utilizzate per la misura
delle traiettorie mandibolari si basano sull'inserimento di magneti sulle arcate
dentarie e la rilevazione delle variazioni del campo elettrico durante i
movimenti (elettrognatografia). Il più delle volte viene analizzato lo
spostamento di un punto incisivo. Il movimento descritto da un punto incisale
inferiore durante un atto masticatorio ha, pur con notevoli variazioni, un
andamento caratteristico. Durante il movimento di apertura, il punto si discosta
di poco dalla linea sagittale mediana o, inizialmente, può spostarsi dal lato
opposto al bolo. Verso la fine del movimento di apertura, si ha uno spostamento
verso il lato di masticazione che continua anche durante la prima metà del
movimento di chiusura, dopo di che si ha un ritorno verso la linea mediana.
Osservato sagittalmente, il movimento di apertura è anteriore rispetto a quello
di chiusura. Il grado di apertura durante l'atto masticatorio varia
notevolmente: circa da 0,8 a 2,2 cm; in ogni caso è massimo all'inizio di una
sequenza masticatoria e tende progressivamente a diminuire. Il massimo
spostamento laterale è di circa 0,8 cm ed è sempre maggiore verso il lato di
masticazione, mentre può mancare del tutto verso il lato opposto. Lo
spostamento laterale verso il lato di masticazione è maggiore con cibi duri che
con cibi molli e tende a diminuire man mano che il cibo viene masticato. La
velocità del movimento è maggiore durante la fase di apertura che in quella di
chiusura (sembra che la velocità di chiusura durante la masticazione naturale
diminuisca rapidamente prima che i denti incontrino la massima resistenza da
parte del bolo: ciò suggerirebbe l'esistenza di un meccanismo di controllo
neuromuscolare che interverrebbe prima che la forza di chiusura sia molto
pronunciata). Un contatto interdentario in massima intercuspidazione è presente
alla fine dell'atto masticatorio nella maggioranza dei casi, e questa posizione
viene mantenuta durante la pausa del movimento che precede l'inizio del
successivo atto masticatorio. Durante una prima fase di questa pausa in ICP,
l'attività dei muscoli di chiusura permane elevata. Vi è tuttavia un breve
periodo silente subito dopo il contatto dentario. Contatti interdentari al di
fuori della posizione di massima intercuspidazione intervengono in vari casi
nella fase finale del movimento di chiusura e, più raramente e per
un'estensione più breve, nella fase iniziale della apertura. La maggioranza dei
soggetti non presenta un ciclo masticatorio simmetrico sui due lati, ma ha un
lato di masticazione preferenziale.
Durante la fase di apertura, i due condili si spostano fin dall'inizio in basso
ed in avanti: nei movimenti funzionali non è presente infatti un movimento
rotatorio a cerniera. Si portano quindi lateralmente (il condilo lavorante) e
medialmente (il condilo bilanciante) e ritornano indietro durante il movimento
di chiusura. In questa fase il percorso di ritorno del condilo lavorante si
colloca inferiormente rispetto al percorso di andata durante l'apertura. Nella
fase terminale di chiusura, in cui vi è per lo più un contatto di scorrimento
tra i denti antagonisti del lato lavorante, il condilo lavorante (che si trova
in posizione laterale e retrusa) si porta in avanti e medialmente, in
intercuspidazione. Questo movimento è l'inverso di quello precedente, descritto
come "spostamento laterale immediato". Per quanto riguarda la sua
ampiezza, ha un valore medio di 0,18 mm di componente mediale e di 0,3 mm di
componenete anteriore. Nello stesso momento il condilo bilanciante si porta in
alto, indietro e lateralmente, nella sua posizione di intercuspidazione. Il
primo molare lavorante si sposta, in quest'ultima fase, medialmente, in alto ed
in avanti in intercuspidazione, mentre il primo molare bilanciante si sposta in
alto e lateralmente, ma non ha una componente di movimento anteriore.
Nella fase finale di un ciclo masticatorio, le superfici occlusali dei denti
antagonisti entrano in contatto. Il contatto avviene dapprima tra i molari del
lato di bilanciamento (opposto a quello dove si trova il bolo) e poi tra i
molari del lato di lavoro (dove si trova il bolo). In tale situazione, quindi,
la mandibola si inclina dapprima facendo perno sul bolo e poi per il contatto
tra i denti del lato di bilanciamento. In questa fase il legamento periodontale
può esercitare una funzione di cuscinetto consentendo al dente un movimento
assiale, non superiore a 0,05 mm.
MASTICAZIONE
(Terza parte)
Fattori muscolari
Muscoli masticatori. La
mobilità della mandibola nella masticazione e nella fonazione si deve ad un
gruppo di muscoli del cranio e del collo, per le cui caratteristiche e dettagli
sulle sedi di inserzione si rimanda ai più opportuni trattati di anatomia.
I muscoli della masticazione vengono normalmente distinti in elevatori ed
abbassatori della mandibola. Gli elevatori sono il massetere, lo pterigoideo
interno (mediale) ed il temporale. I primi due hanno un decorso grossolanamente
parallelo in basso ed in dietro: il massetere decorre con due strati,
superficiale e profondo, dall'arcata zigomatica alla faccia laterale dell'angolo
e del corpo della mandibola; lo pterigoideo interno va dalla fossa pterigoidea
dello sfenoide alla faccia mediale dell'angolo della mandibola. Il temporale
decorre dalla linea temporale inferiore e dalla fossa temporale al processo
coronoideo della mandibola: vi si distinguono fasci anteriori, medi e
posteriori. I primi hanno un andamento verticale, gli altri obliquo o in basso
ed in avanti. Mentre i muscoli massetere e pterigoideo interno sono
essenzialmente muscoli di elevazione pura, il muscolo temporale, con i suoi
fasci medi e posteriori, è responsabile della componente retrusiva dei
movimenti mandibolari (ritorno del condilo all'indietro nella cavità glenoidea
durante i movimenti di chiusura della bocca). La sola contrazione dei fasci
posteriori fa compiere alla mandibola movimenti di retrusione pura. Nei
movimenti di chiusura della bocca si contrae anche il capo superiore del muscolo
pterigoideo esterno, che decorre dal menisco articolare al piano infratemporale
dello sfenoide: in tal modo viene controbilanciata la tensione delle fibre
elastiche dell'inserzione meniscale posteriore.
I muscoli abbassatori della mandibola sono essenzialmente: il miloioideo, il
genioioideo ed il digastrico. I fasci del muscolo miloioideo decorrono dalla
linea miloioidea della mandibola in basso, in dietro e medialmente per inserirsi
su di un rafe mediano, mentre i fasci più posteriori si fissano sulla faccia
anteriore del corpo dell'osso ioide. Il muscolo genioiodeo si poggia sulla
faccia superiore del muscolo miloioideo e va dalla spina mentale alla faccia
anteriore del corpo dell'osso ioide. Il muscolo digastrico presenta due ventri,
anteriore e posteriore, con un tendine intermedio. Il ventre posteriore inizia
dall'incisura mastoidea del temporale e si continua, in basso ed in avanti, nel
tendine intermedio che è collegato all'osso ioide per mezzo di un'ansa fibrosa.
Il ventre anteriore si porta dal tendine intermedio alla fossetta digastrica
della mandibola. Questi tre muscoli abbassano la mandibola se l'osso ioide è
fisso (masticazione), mentre innalzano lo ioide se è fissa la mandibola
(deglutizione). All'apertura della bocca partecipa anche il capo inferiore del
muscolo pterigoideo esterno, che si inserisce al collo del condilo ed al
processo pterigoideo dello sfenoide. La contrazione singola unilaterale del capo
inferiore dello pterigoideo esterno sposta la mandibola verso il lato opposto;
la contrazione bilaterale la porta in avanti.
La sequenza di azione dei vari muscoli durante la masticazione è molto
complessa, con sovrapposizioni dell'attività di muscoli diversi, anche
antagonisti. Durante le fasi di apertura del movimento masticatorio viene
attivato prima il muscolo miloioideo, poi il digastrico e poi il capo inferiore
dello pterigoideo esterno. L'inizio di attività del digastrico coincide però
con il momento di massima attività del fascio anteriore del muscolo temporale,
durante la fase terminale della precedente chiusura. Raffrontando l'attività
muscolare, valutata elettro-miograficamente, con i vari gradi di forza di
chiusura, misurata interponendo tra i denti un trasduttore e chiedendo al
soggetto di esercitare una pressione dentaria progressivamente crescente, si
vede che, superato un certo grado di pressione, si ha un'attivazione dei muscoli
abbassatori insiema a quella, già presente, degli elevatori. Ciò può spiegare
la sovrapposizione dell'attività del muscolo digastrico al momento della
massima attività del temporale anteriore che si osserva alla fine del movimento
masticatorio. Durante la fase di chiusura, il primo muscolo ad essere attivato
è lo pterigoideo interno, che precede di 10 - 40 msec gli altri muscoli
elevatori (temporale anteriore, temporale posteriore e massetere). Nel corso
della masticazione unilaterale, l'attività del massetere mostra un andamento
caratteristico: l'attività del muscolo dal lato opposto a quello di
masticazione precede quella del muscolo dello stesso lato e diminuisce
lentamente dopo aver raggiunto il massimo di tensione; il muscolo dello stesso
lato presenta un'attività più intensa, con brusca riduzione dopo aver
raggiunto il massimo. Durante la fase di chiusura viene attivato anche il capo
superiore dello pterigoideo esterno.
Esiste una grande varietà da soggetto a soggetto per numero di movimenti
masticatori e ampiezza delle contrazioni muscolari. L'intensità della
contrazione è normalmente maggiore dal lato di lavoro: è così quasi sempre
agevole determinare il lato di masticazione durante un ciclo di masticazione
spontanea. Il rapporto di intensità di contrazione è spesso all'incirca di 2:1
a favore del lato di lavoro, ma può variare notevolmente nei diversi soggetti.
Individui sani con arcate dentarie integre possono presentare un'attività
muscolare quasi identica sui due lati. Durante il ciclo masticatorio, il bolo
viene per lo più trasferito due o tre volte da un lato all'altro, ma è
frequente osservare la presenza di un lato dominante, in cui la masticazione
avviene per un periodo di tempo più prolungato.
Labbra, guance e lingua. Le
labbra sono due ripiegature muscolo-mucose che costituiscono la parete anteriore
del vestibolo della bocca e delimitano l'orifizio buccale. Applicate alle arcate
alveolo-dentarie, ne ripetono la forma e, generalmente, corrispondono in altezza
a queste ultime. Le labbra sono indispensabili per l'assunzione degli alimenti
nel lattante (v. deglutizione) e mantengono questa loro funzione nella vita
adulta, quando gli alimenti liquidi si assumono per sorbimento. Occorre allora
che le labbra siano applicate all'orlo del bicchiere o alla superficie del
liquido, creando poi una pressione negativa nel cavo orale. La partecipazione al
linguaggio parlato (v. fonazione) fa delle labbra un organo indispensabile alla
pronuncia dei fonemi cosiddetti labiali, ma non é trascurabile il contributo
che esse portano alla formazione di tutti i suoni. Si parla di inadeguatezza
delle labbra se la conformazione dei tessuti molli é tale da non consentire la
chiusura della rima labiale quando la mandibola é nella posizione di riposo ed
i muscoli mimici sono rilassati. Non sempre, comunque, una morfologia labiale
adeguata è sufficiente a garantire il sigillo anteriore. In tutti quei casi, ad
esempio, in cui vi sia una ostruzione delle vie nasali (per ingrossamento delle
adenoidi, riniti, deviazioni del setto nasale, ecc.) il soggetto é costretto a
respirare con la bocca, e l'adeguatezza viene meno. In questi casi il sigillo
anteriore viene recuperato con la rimozione della causa ostruente ed il
ripristino della funzione nasale.
Le guance, come le labbra, sono pareti muscolo-mucose rivestite all'esterno da
cute ricca di follicoli piliferi e ghiandole sebacee, che ricoprono le regioni
zigomatica, masseterina e geniena. Quest'ultima rappresenta la parte mobile
delle guance ed è costituita in massima parte dai muscoli buccinatore e
massetere. Oltre alla masticazione, contribuiscono alla genesi delle variazioni
pressorie nella cavità orale e coadiuvano la lingua nel mantenere il bolo
alimentare sotto le arcate dentarie, durante la masticazione.
Sia le labbra che le guance partecipano alla mimica del volto ed influenzano la
direzione degli assi dentari. A tale proposito, il contributo delle labbra è
superiore a quello delle guance e pari, forse, a quello della lingua. L'azione
di quest'ultima, che tende alla esteriorizzazione dei denti anteriori, é
normalmente bilanciata da quella delle labbra, che si esercita in direzione
opposta.
La lingua é un organo muscolare che adempie ad importanti funzioni in rapporto
con la masticazione, la deglutizione e la fonazione. Le formazioni muscolari,
che ne costituiscono la massima parte, provengono dalla mandibola e dall'osso
ioide e, sporgendo sul pavimento della bocca, ne vengono rivestite dalla mucosa.
La lingua é inoltre importante sede della funzione del gusto, per la presenza
delle papille gustative. La regione della punta é anche dotata di elevata
sensibilià tattile, con potere discriminatorio (acuità tattile) intorno ad 1
mm.
Retraendosi posteriormente, mentre le labbra sono sigillate al capezzolo o ad un
contenitore, la lingua provoca la depressione necessaria per la suzione. Durante
la masticazione, distribuisce e mantiene gli alimenti sulla superficie
masticatoria dei denti. Molto importante é la sua funzione nel primo tempo
della deglutizione (volontario), in quanto contribuisce a spingere il bolo nella
faringe attraverso le fauci. Altra funzione fondamentale della lingua é svolta
nel linguaggio articolato. Se esistono, infatti, fonemi chiamati linguali per la
cui formazione la lingua é essenziale, tutti i fonemi sarebbero impossibili o
incomprensibili senza di essa.
Pur essendo costituita da muscolatura striata, l'azione della lingua é
principalmente incosciente, guidata dai suoi recettori tattili e da quelli
presenti in tutta la mucosa orale, ovvero da schemi motori appresi durante
l'infanzia e successivamente difficilmente modificabili. La lingua infatti, come
detto, é un organo tattile finemente discriminativo, ma invia stimoli in gran
parte solo al livello tronco-encefalico, e scarse informazioni raggiungono il
livello cosciente (abbastanza grossolana é la consapevolezza della sua
posizione nella cavità orale). Durante l'accrescimento, infine, la lingua
svolge una funzione determinante sull'evoluzione della architettura ossea della
mandibola, dei mascellari e delle altre componenti dello splancnocranio, come
pure influenza la direzione degli assi dentari.
Fattori nervosi
La masticazione è un evento ritmico, la cui sequenza è generata nel tronco
dell'encefalo, che viene modificato da informazioni provenienti dal sistema
nervoso centrale e da quello periferico. Non la sequenza degli atti masticatori,
ma il loro ritmo sarebbe invece determinato da un centro sito nella sostanza
reticolare della base dell'encefalo. Ad un livello più basso di organizzazione
gerarchica, sono coinvolti i nuclei motori dei nervi cranici ed i nuclei
sensitivi, specialmente trigeminale e tratto solitario. Questi ultimi ricevono
informazioni dalla periferia e le trasmettono ai centri superiori oppure,
direttamente o indirettamente, ai motoneuroni dei nervi cranici, come avviene in
alcuni riflessi. Il ruolo della corteccia cerebrale nella masticazione non è
ancora completamente chiarito. La zona motrice corticale per la lingua e la
mandibola è localizzata nei giri precentrale e postcentrale. Nella scimmia, la
zona corticale motrice dei singoli muscoli si modifica variando la profondità
della stimolazione corticale. E' probabile che la funzione della corteccia
cerebrale sia quella di iniziare il movimento di apertura della bocca,
all'inizio dell'atto masticatorio, e di elaborare ed integrare le informazioni
provenienti dalla periferia. La corteccia cerebrale, cioé, utilizzando le
informazioni che le giungono dal cavo orale e dall'apparato muscolo-articolare
della mandibola, può modificare, nel corso della masticazione, sia l'attività
stessa del generatore centrale che i riflessi che controllano i muscoli
masticatori. Nella scimmia, l'ablazione bilaterale dell'area masticatoria,
dell'area cioè la cui stimolazione produce movimenti ritmici mandibolari, é
seguita da gravi difficoltà ed alterazioni della masticazione, particolarmente
accentuate nel primo periodo post-operatorio. La lesione provoca anche
alterazioni gravi del movimento della lingua e della deglutizione. Nel
successivo periodo, l'animale riprende a masticare, ma spesso permangono a
lungo, o definitivamente, alterazioni del normale ritmo masticatorio.
L'alterazione residua è tale da suggerire che, in mancanza dell'area corticale
motrice, il generatore sottocorticale possa semplicemente determinare
un'alternanza nell'attività dei motoneuroni trigeminali antagonisti, con
ridotte possibilità di modificarne in modo dinamico l'attività, adattandola
alle mutate condizioni della periferia. Si possono pertanto ipotizzare almeno
due vie efferenti dalla corteccia cerebrale: la prima diretta al nucleo motore
del trigemino e la seconda al centro della masticazione nel tronco
dell'encefalo. Il ritmo masticatorio sarebbe invece sostenuto da un altro
centro, situato nella sostanza reticolare.
Innervazione sensitiva del territorio oro-faciale.
Le informazioni esterocettive e propriocettive originate dal cavo orale e dalle
strutture ad esso annesse vengono utilizzate per i processi della masticazione e
della deglutizione ed anche per l'articolazione della parola.
Le fibre afferenti primarie della sensibilità somatica che innervano la cute e
gran parte delle strutture profonde della faccia (muscoli, ossa ed
articolazioni), le mucose della cavità orale e di quella nasale, ed i denti
fanno capo prevalentemente al nervo trigemino (V nervo cranico). Tali fibre
entrano nel ganglio semilunare di Gasser attraverso le tre branche o nervi
principali del trigemino: oftalmica, mascellare e mandibolare. Il nervo
oftalmico innerva la cute della porzione anteriore del cuoio capelluto, della
fronte e del naso nonchè la cornea, la congiuntiva e la mucosa dei seni
frontale, sfenoidale e del setto nasale. Il nervo mascellare innerva la cute
della porzione intermedia della faccia ed il labbro superiore, le mucose del
seno mascellare, della cavità nasale, del rinofaringe, della metà superiore
della cavità orale ed i denti dell'arcata superiore. Il nervo mandibolare
innerva la cute della metà inferiore della faccia, la mucosa della metà
inferiore del cavo orale, i 2/3 anteriori della lingua e l'arcata dentaria
inferiore. Tutte le fibre hanno le cellule di origine nel ganglio semilunare, ad
eccezione delle fibre originate dai fusi neuro-muscolari dei muscoli masticatori
e di alcuni meccanocettori del legamento periodontale che attraversano il
ganglio semilunare, avendo la cellula di origine nel nucleo mesencefalico del
trigemino che, da questo punto di vista, ha caratteristiche assolutamente
uniche, in quanto rappresenta un vero e proprio ganglio radicolare inserito nel
sistema nervoso centrale. Altre fibre afferenti decorrono nel nervo faciale (VII),
nel glosso-faringeo (IX) e nel vago (X). Nel nervo faciale decorrono fibre
provenienti dal padiglione auricolare; nel nervo glosso-faringeo fibre originate
dal palato molle, dall'istmo delle fauci, dalla faringe e dal terzo posteriore
della lingua, nonchè dall'orecchio medio e dalla tuba uditiva; nel nervo vago
decorrono fibre originate dalla faccia mediale del padiglione auricolare e dal
meato acustico. Le afferenze somatiche che decorrono in questi nervi entrano a
far parte del tratto spinale del trigemino e terminano nei nuclei di relay
trigeminale.
Tutte le classi di recettori somatici (tatto, pressione, temperatura, dolore)
sono rappresentate nel territorio intraorale. L'attivazione di questi recettori
provoca effetti sinaptici complessi sui motoneuroni trigeminali che innervano i
muscoli masticatori. La stimolazione delle fibre afferenti contenute nei diversi
nervi sopra menzionati ha sempre effetti eccitatori sui motoneuroni dei muscoli
abbassatori. L'effetto è mediato da almeno due sinapsi, una a livello degli
stessi motoneuroni, l'altra nel complesso nucleare trigeminale. Quanto detto è
valido anche per i recettori parodontali. Per quanto riguarda i muscoli
elevatori, si sono osservati, invece, sia effetti inibitori che effetti
eccitatori. La stessa stimolazione che eccita i motoneuroni degli abbassatori
inibisce i motoneuroni dei muscoli elevatori.
Gli effetti inibitori consistono nella comparsa di una iperpolarizzazione
precoce seguita da una tardiva. Il primo effetto è mediato da una via
disinaptica, il secondo ha invece maggior latenza ed è mediato da una via
polisinaptica. Le due vie sono indipendenti l'una dall'altra ed includono gruppi
distinti di interneuroni inibitori. Il potenziale inibitorio di breve latenza,
disinaptico, è mediato da interneuroni situati nel nucleo infratrigeminale e
sopratrigeminale di Lorente de Nò, costituito da un aggregato di piccole
cellule che circondano dorsalmente e medialmente il nucleo motorio del
trigemino. Da questi interneuroni originano anche fibre commissurali che si
portano al nucleo motorio del trigemino contralaterale, spiegando l'inibizione
bilaterale che si osserva per stimolazione dei nervi di un solo lato (inibizione
riflessa crociata). Il potenziale inibitorio di lunga latenza, invece, è
mediato da neuroni situati a livello del nucleo orale-interpolare, che a sua
volta proietta ai neuroni della formazione reticolare bulbare, probabilmente i
veri responsabili della generazione del fenomeno.
La stimolazione delle fibre a più bassa soglia contenute nei nervi intraorali
(comprese quelle che originano dal legamento periodontale) provoca anche effetti
eccitatori sui motoneruroni dei muscoli elevatori.
toressa
Recettori dell'articolazione temporo-mandibolare.
Questi recettori hanno un ruolo importante nella percezione cosciente del
movimento e della posizione della mandibola e probabilmente partecipano anche
alla regolazione riflessa del movimento mandibolare. Nella capsula e nei
legamenti della ATM sono stati identificati istologicamente quattro tipi di
recettori: recettori di Ruffini, di Golgi, di Pacini e terminazioni libere di
fibre mieliniche ed amieliniche, molte delle quali di tipo nocicettivo. I
recettori sono più numerosi nelle regioni laterali e posteriori della capsula
articolare e sono innervati da fibre che decorrono nel nervo auricolo-temporale.
Sono stati identificati due tipi funzionali di recettori: a rapido ed a lento
adattamento. I primi si eccitano solo all'inizio di un movimento di
abbassamento, ovvero all'inizio di un movimento di elevazione della mandibola.
L'eccitazione è determinata dalla rapida deformazione della capsula articolare,
provocata dalla rotazione del condilo. I recettori a lento adattamento scaricano
invece per tutto il tempo in cui il condilo è ruotato, ma non nella posizione
di riposo; la frequenza di scarica è in funzione della estensione del movimento
articolare.
Gli effetti centrali provocati dalla stimolazione dei recettori della ATM sono
poco noti. Parte di questi recettori, almeno quelli eccitati dal normale
movimento dei condili, provoca effetti eccitatori sui muscoli elevatori quando
la mandibola viene abbassata.
Recettori muscolari e tendinei.
Appartengono
a questa classe di recettori i fusi neuromuscolari e gli organi tendinei del
Golgi, identici, per morfologia e funzioni, a quelli degli altri distretti
corporei. La distribuzione di questi recettori nell'apparato masticatorio non
è, però, omogenea come altrove. I muscoli elevatori (antigravitari) contengono
un elevato numero di recettori fusali che non sono stati dimostrati, invece, nei
muscoli abbassatori. Nonostante siano stati occasionalmente descritti recettori
tendinei del Golgi nel muscolo massetere, è opinione corrente che tali
recettori, od altri recettori funzionalmente analoghi, siano del tutto assenti
nei muscoli dell'apparato masticatorio.
Le fibre originate dai fusi neuromuscolari formano contatti sinaptici di tipo
eccitatorio con i motoneuroni alfa dei muscoli elevatori ipsilaterali, situati
nel nucleo motorio del trigemino, sia omonimi che eteronimi sinergici. Non si
sono mai osservati effetti inibitori sui motoneuroni dei muscoli abbassatori.
Pertanto, a livello del nucleo motore del trigemino, non vige il principio della
innervazione reciproca, come invece si verifica a livello spinale.
Le afferenze originate dai recettori extra-fusali dei muscoli masticatori
(recettori del Pacini e terminazioni nervose libere), parte dei quali di tipo
nocicettivo, fanno capo alle cellule del ganglio semilunare e proiettano ai
nuclei sensitivi del complesso trigeminale. La loro stimolazione provoca effetti
bilaterali di tipo inibitorio nei motoneuroni dei muscoli elevatori e di tipo
eccitatorio nei motoneuroni degli abbassatori.
Innervazione intradentale.
Le fibre nervose
entrano nella cavità dentaria attraverso il foro radicolare e si irradiano
verso la zona parietale della polpa, formando un plesso nervoso al di sotto
della zona ricca di cellule; si dirigono quindi perifericamente per raggiungere,
non tutte, lo strato degli odontoblasti, inserendosi tra di loro. A questo
livello le fibre emettono numerose collaterali, ed in prossimità della
superficie dentinica formano il plesso marginale. Molte fibre entrano e
terminano nei tubuli dentinali, affiancando i processi odontoblastici.
Alcune fibre afferenti della polpa dentaria possono, prima di entrare nel foro
apicale, dividersi in una o più branche ed innervare la gengiva o il periostio
in prossimità del dente, o perfino un dente limitrofo.
Le fibre intradentali sono eccitabili da una vasta gamma di stimoli applicati
alla dentina. Stimoli meccanici, chimici e termici sono in grado di provocare
eccitazione delle fibre intradentali. Gli stimoli termici applicati al dente
devono, per essere efficaci, superare i 50 °C o essere inferiori ai 17 °C. Gli
stimoli termici e gran parte di quelli chimici provocano dolore se applicati
alla dentina o direttamente nella polpa dentaria. Non è chiaro come gli stimoli
naturali possano attivare le fibre intradentali. I rapporti morfologici tra gli
odontoblasti ed i terminali nervosi, comunque, sono tali da far ritenere che le
due strutture siano fra loro funzionalmente accoppiate. Sono state formulate due
diverse ipotesi, che prevedono rispettivamente un accoppiamento di tipo
sinaptico o elettrico ed uno di tipo meccanico.
Secondo la prima ipotesi, l'odontoblasta funzionerebbe da cellula recettrice e
svilupperebbe, stimolato, un potenziale generatore che, per effetto di
liberazione di sostanze chimiche o per accoppiamento elettrotonico,
provocherebbe a sua volta depolarizzazione ed eccitazione dei terminali delle
fibre afferenti. Non esistono prove dirette a convalidare tale ipotesi.
La seconda ipotesi si basa su un meccanismo idrodinamico, abbastanta verosimile
ed osservato nei tubuli dentinali in vitro. Gli stimoli applicati alla dentina
provocherebbero una deformazione dei processi odontoblastici che verrebbe
meccanicamente trasmessa ai terminali nervosi situati più profondamente nei
tubuli o tra i corpi delle cellule odontoblastiche. L'insorgenza dell'impulso
elettrico nel terminale assonico avverrebbe quindi per deformazione meccanica
della sua membrana, in modo simile a quanto avviene in altri territori somatici.
Quale che sia la natura dello stimolo applicato alla dentina o alla giunzione
amelo-dentinale, la sensazione riferita è comundue di dolore. Ciò si accorda
con l'ipotesi del meccanismo idrodinamico della trasduzione degli stimoli,
secondo cui qualsiasi stimolo capace di promuovere movimento di fluidi
nell'interno dei tubuli dentinali, (o che comunque provochi dilatazione dei
processi odontoblastici) eccita i terminali nervosi e provoca dolore.
Innervazione del legamento periodontale.
Il
legamento periodontale è riccamente innervato da grosse fibre mieliniche ed
amieliniche. Parte delle fibre amieliniche è di tipo vegetativo, associata ai
piccoli vasi del periodonzio ed in parte origina da nocicettori. Le fibre
mieliniche di grosso diametro terminano in corpuscoli ed apparati recettoriali
più o meno complessi, che potrebbero svolgere la funzione di meccanocettori.
Questi recettori dovrebbero essere considerati veri e propri recettori profondi
o propriocettori, essendo situati in un tessuto articolare, ma vengono in genere
inclusi fra i pressocettori del cavo orale. Possono anche essere considerati un
tipo speciale di recettori di tatto-pressione, in quanto responsabili della
sensazione coscente di contatto del dente. Talune fibre afferenti dei
meccanocettori periodontali hanno la cellula di origine nel ganglio semilunare,
altre nel nucleo mesencefalico del trigemino. Il movimento del dente, sottoposto
a pressioni esercitate nelle diverse direzioni, provoca compressioni in alcune
zone dei tessuti periodontali e trazioni in altre. E' probabile che taluni
recettori vengano eccitati sia dalla compressione che dalla trazione, mentre
altri sono eccitati dall'una ed inibiti dall'altra. La soglia di eccitabilità
dei meccanocettori periodontali è molto bassa: sono in genere sufficienti
spostamenti del dente di soli 2 micron. La soglia, espressa in grammi, è
nell'ordine di 3 - 10 g, ma alcune fibre possono scaricare per un carico di un
solo grammo. La soglia più bassa è stata trovata nei meccanocettori degli
incisivi, che hanno la maggiore mobilità, mentre la più alta per quelli dei
molari, che sono provvisti di un sistema più rigido di ancoraggio all'alveolo.
I meccanocettori periodontali possono essere sia a rapido che a lento
adattamento. Le unità a rapido adattamento sono sensibili alla velocità dello
spostamento, quelle a lento adattamento sono invece egualmente sensibili alla
velocità (fase dinamica) che alla dislocazione statica del dente, in quanto
rimangono eccitate per tutto il tempo in cui la forza viene mantenuta costante
(fase statica). La frequenza di scarica dei recettori a lento adattamento è
proporzionale, entro certi limiti, alla intensità della forza applicata.
Praticamente tutti i recettori periodontali rispondono in modo differenziato a
forze applicate al dente in una particolare direzione, anche se esiste una
soglia più bassa per stimoli applicati in una direzione ottimale.
L'intera categoria dei meccanocettori periodontali ha azioni complesse sui
motoneuroni del trigemino, potendo sia eccitare che inibire i motoneuroni dei
muscoli elevatori. E' probabile che i tipi di recettori che provocano effetti
sinaptici di segno opposto siano tra loro diversi, o per soglia o per
sensibilità a stimoli specifici, ovvero che le loro afferenze vengano
controllate a livello centrale da un meccanismo nervoso che ne può selezionare
l'utilizzazione.
Funzioni riflesse delle afferenze periferiche.
Esistono strette correlazioni funzionali tra strutture di controllo della
posizione delle due arcate dentarie e l'intero apparato muscolare motore della
mandibola. Le pressioni esercitate dai singoli denti, nella chiusura della bocca
e nella masticazione dei cibi, rilevate dai recettori peridontali, della capsula
articolare e dei ligamenti, vengono trasmesse ai muscoli della masticazione.
Essi possono in tal modo adeguare la loro forza di contrazione ed il loro tono
di riposo ai riferimenti provenienti dai denti e dall'articolazione
temporomandibolare. Possono comparire disfunzioni dell'apparato muscolare della
mandibola quando si hanno alterazioni importanti della fisiologica articolarità
dentaria.
Riflesso mandibolare di chiusura e postura della
mandibola.
I riflessi da
stiramento dei muscoli elevatori, gli unici dotati di fusi neuromuscolari,
vengono distinti in fasici e tonici. Il riflesso fasico, noto anche come
riflesso di chiusura della mandibola o riflesso masseterino, è l'equivalente
dei riflessi miotatici o tendinei degli arti. E' evocabile percuotendo
leggermente il mento con un martelletto, in un soggetto che abbia appena aperto
la mandibola ed abbia i muscoli rilassati. L'abbassamento rapido della mandibola
è seguito da una rapida contrazione fasica del massetere e del temporale. Il
riflesso da stiramento tonico è provocato dalle stesse afferenze fusali (specie
quelle originate dalle terminazioni secondarie), quando la mandibola è
mantenuta abbassata.
Ai riflessi di chiusura della mandibola, provocati per apertura della stessa,
possono anche partecipare le afferenze delle ATM. I riflessi miotatici (e
probabilmente anche quelli iniziati dai recettori articolari) contribuiscono a
mantenere nei muscoli elevatori quell'attività tonica di base che caratterizza
la postura della mandibola, cioè la posizione di riposo della mandibola, chiusa
ad un livello tale da lasciare un certo spazio tra gli incisivi superiori e gli
inferiori. I muscoli elevatori sono tipici muscoli antigravitari, che utilizzano
quindi gli stessi meccanismi per il mantenimento del tono posturale in atto
negli antigravitari delle altre parti del corpo. Non tutti i dati sperimentali
sembrano comunque confermare l'ovvia ipotesi che la postura della mandibola sia
mantenuta attivamente per l'azione riflessa delle afferenze fusali. Abolendo
sperimentalmente queste ultime, infatti, (distruzione del nucleo mesencefalico
del trigemino) la postura mandibolare non sembra alterarsi significativamente.
Probabilmente altri riflessi, iniziati dall'articolazione temporomandibolare e
dal cavo orale, possono vicariare e/o cooperare con quelli miotatici, ovvero la
postura mandibolare potrebbe non essere dovuta a fenomeni riflessi. Si sostiene,
infatti, che la postura della mandibola possa essere in realtà mantenuta ad
opera di soli fenomeni passivi, quali l'elasticità dei muscoli, delle fasce e
dei legamenti, come pure dalla presenza di una pressione intraorale
sub-atmosferica. Probabilmente tutti i sopra citati meccanismi concorrono, in
gradi e momenti diversi, al mantenimento della postura mandibolare. Certamente
il fattore contrazione, attivato per via riflessa, promuove periodicamente quei
necessari aggiustamenti posturali della mandibola che si rendono necessari
quando questa viene sottoposta a forze inerziali che ne inducono l'abbassamento
(variazioni rapide della posizione del capo, fasi di appoggio della marcia o
della corsa).
Altri riflessi di chiusura della mandibola.
La stimolazione di talune afferenze intraorali a bassa soglia, fra cui quelle
originate dai recettori periodontali, può provocare un riflesso di chiusura
della mandibola per contrazione dei muscoli elevatori. La stimolazione
fisiologica dei recettori periodontali non provoca, in realtà, evidenti effetti
riflessi sui muscoli elevatori se questi sono rilassati, ma ne evoca la risposta
contrattile se questi sono in attività, come si ha mordendo un oggetto
interposto tra le arcate dentarie: riflesso
periodonto-masseterico.
Riflessi di apertura della mandibola.
Per
lungo tempo si è ritenuto che la stimolazione delle afferenze intraorali
provocasse solo riflessi di apertura della mandibola. In realtà ciò è vero
solo per le fibre afferenti che veicolano informazioni di natura dolorifica. La
registrazione contemporanea dell'attività del massetere e del digastrico
evidenzia che il primo viene inibito ed il secondo eccitato, indicando così la
presenza di un'organizzazione centrale del riflesso nocicettivo analoga a quanto
presente nelle altre parti dell'organismo (riflessi flessori). Il significato
funzionale dei riflessi di apertura è palesemente protettivo nei confronti
della integrità dei tessuti intraorali.
Compensazione riflessa della contrazione da variazione
di carico.
Esistono due situazioni, tra loro opposte, che possono
verificarsi in un normale ciclo masticatorio e che esemplificano, in parte,
quanto esposto circa i riflessi mandibolari: un cibo friabile, che offre
inizialmente una certa resistenza, cede rapidamente alla pressione esercitata
fra le arcate dentarie, oppure un cibo, inizialmente tenero, offre un
inaspettata resistenza.
Nel primo caso, a seguito del cedimento del bolo, si verifica una risposta
riflessa che comporta la rapida decontrazione del massetere e la comparsa di
attività nel muscolo digastrico, impedendo l'occlusione violenta delle arcate
dentarie. La soppressione dell'attività del muscolo massetere avviene in due
fasi: una rapida (dopo circa 8 msec) ed una tardiva (dopo circa 30 msec), mentre
per il digastrico esiste solo la componente tardiva (circa 30 msec). La risposta
rapida è verosimilmente mediata da un arco riflesso breve (a partenza
probabilmente dalle terminazioni fusali, rapidamente detese dall'accorciamento
del muscolo), mentre quella tardiva potrebbe coinvolgere anche la corteccia
cerebrale.
Nel caso opposto, l'improvvisa applicazione di un carico è seguita, dopo circa
8 - 10 msec, da un rapido incremento della contrazione del massetere. L'effetto
potrebbe essere dovuto ad un brusco incremento della scarica fusale: durante la
cotrazione, infatti, la scarica nelle efferenti gamma aumenta progressivamente
per adattare la lunghezza dei recettori al progressivo accorciamento del
muscolo; quando l'accorciamento del muscolo viene impedito, la scarica gamma
continua, promuovendo uno stiramento delle terminazioni anulospirali
relativamente maggiore a quanto dovuto per l'accorciamento delle fibre muscolari
extrafusali e scatenando, quindi, un riflesso miotatico.
Controllo centrale della masticazione.
Il meccanismo della normale masticazione è più complesso della semplice
alternanza di riflessi di apertura e chiusura della mandibola, scatenati dalla
presenza di contenuto nella cavità orale e dalla tensione dei muscoli
masticatori. Per via riflessa possono essere provocati solo semplici movimenti
di apertura e di chiusura, ma non la successioni di eventi masticatori che si ha
in una normale sequenza, in condizioni fisiologiche. La masticazione è invece
iniziata e mantenuta dall'attività di centri troncoencefalici che possono
operare anche indipendentemente dalle afferenze periferiche. Questi centri sono
chiamati, collettivamente, generatore centrale. Per la programmazione di un
determinato tipo di masticazione, questo si avvale delle informazioni
provenienti dalla periferia, può essere modulato e può a sua volta modulare
l'azione degli archi riflessi. Il generatore centrale è, a sua volta, sotto il
controllo di livelli superiori, corticali e sottocorticali. Le afferenze
periferiche forniscono ai vari livelli le informazioni loro necessarie e
modulano l'attività riflessa dei motoneuroni trigeminali. Il modello di base
del ritmo masticatorio (alternanza di aperture e chiusure) è generato
dall'attività nervosa centrale, ma il tipo, l'intensità, la durata e la
frequenza dei movimenti dipendono dalle caratterisitche del cibo. I cicli della
masticazione, quindi, e le forze prodotte dai muscoli che li promuovono, sono
tral loro diversi e sempre adatti alla situazione contingente, infatti le
afferenze periferiche sono di fondamentale importanza per programmare
l'attività del generatore centrale all'introduzione del cibo nella cavità
orale e per modificarla mano a mano che la masticazione procede.
DEGLUTIZIONE
GUSTO
La sensibilità gustativa fornisce informazioni su alcune caratteristiche
chimiche delle sostanze che vengono a contatto con le mucose dell' orofaringe. I
recettori gustativi (cellule gustative primarie) interagiscono con le molecole
disciolte nella saliva e trasducono lo stimolo chimico in segnali nervosi. Il
sapore è una caratteristica dipendente dalla composizione della sostanza. Si
ritiene che le diverse centinaia di sapori percepibili derivino dalla
combinazione di alcune qualità primarie: dolce, salato, amaro e acido. I sapori
primari differiscono per la natura chimica dei composti in grado di evocarli e
per i meccanismi di captazione e di trasduzione. Contemporaneamente alla
stimolazione gustativa, molecole volatili liberate dalla sostanza possono
legarsi ai recettori della mucosa olfattiva. La sensazione globale che ne deriva
è chiamata aroma.
L'importanza della sensibilità gustativa è collegata soprattutto alla
nutrizione e all'assunzione di liquidi. Il sapore di un cibo o di una bevanda è
infatti fondamentale nel determinarne la commestibilità e l'appetibilità; il
rifiuto di un cibo a causa del suo sapore sgradevole è importante per la
sopravvivenza di molte specie animali. Schematicamente, si possono associare ai
4 sapori fondamentali funzioni diverse: assicurare le riserve energetiche
(dolce), mantenere l'equilibrio elettrolitico (salato), monitorare il pH (acido,
amaro), evitare sostanze tossiche (amaro). Di norma, le sostanze dolci provocano
una sensazione piacevole e l'innesco di riflessi di salivazione, di deglutizione
e di preparazione del tubo digerente alla digestione e all'assorbimento. Invece,
il sapore amaro provoca sovente il rifiuto del cibo o della bevanda e, se molto
intenso, stimola il riflesso del vomito: questo è presumibilmente in relazione
al fatto che la maggior parte delle sostanze tossiche presenti in natura ha
sapore amaro.
Il grado di piacevolezza di un sapore è tuttavia soggettivo e può essere
influenzato dall'esperienza e dalle necessità nutrizionali. Sia l'animale che
l'uomo tendono a rifiutare un cibo dal quale in passato siano stati intossicati.
In base alle informazioni gustative e olfattive si ha la capacità di operare
una scelta tra diversi cibi e preferire quello che fornisce l'apporto nutritivo
più consono alle esigenze dell'organismo.
Meccanismi e vie gustative
Struttura dell'apparato recettoriale.
I
recettori gustativi sono cellule epiteliali specializzate, organizzate in
strutture dette bottoni gustativi.
I bottoni gustativi dei mammiferi
contengono alcune decine di recettori, oltre ad elementi cellulari immaturi
(cellule basali) La porzione apicale, dotata di microvilli, delle cellule
gustative primarie entra in contatto con la saliva che bagna la superficie
epiteliale mediante un canalicolo (poro gustativo). Tramite la porzione basale,
i recettori stabiliscono contatti sinaptici di tipo chimico con le terminazioni
di fibre afferenti primarie. Ogni fibra innerva numerosi bottoni anche in zone
dell'epitelio linguale molto distanti tra loro e, nello stesso bottone, diversi
recettori. Allo stesso tempo, ogni recettore gustativo è innervato da più
fibre afferenti. Morfologicamente, si possono distinguere diversi tipi di
recettori gustativi, cui corrispondono verosimilmente differenze di tipo
funzionale, ancora non chiarite. Le cellule basali costituiscono una popolazione
di cellule staminali che garantisce il ricambio dei recettori gustativi (il
ciclo vitale delle cellule gustative mature è di circa dieci giorni, nei
mammiferi).
I bottoni gustativi sono largamente diffusi nell'epitelio della lingua,
particolarmente nella sua porzione dorsale, ma sono presenti anche nella mucosa
del palato, della faringe, dell'epiglottide e del terzo superiore dell'esofago.
Nella lingua, sono per lo più localizzati nelle papille, strutture
specializzate di cui si distinguono diversi tipi: nell'uomo, solo le papille
vallate, fogliate e fungiformi hanno bottoni gustativi. Ognuna delle papille
vallate e fogliate contiene migliaia di bottoni. Le papille vallate sono
concentrate nella parte posteriore del dorso della lingua, a livello della
'V" linguale. Le papille fogliate sono maggiormente presenti sui lati della
lingua. Le papille fungiformi si trovano numerose sulla punta della lingua e
contengono pochi bottoni gustativi: da I a 12 per papilla.
|
|
| A) Tipi di papille linguali. Le
papille fungiformi, foliate e vallate contengono bottoni gustativi. B)
Distribuzione delle aree linguali di maggior sensibilità |
Trasduzione degli stimoli gustativi.
La superficie recettoriale non risponde in modo omogeneo ai sapori primari. La
punta della lingua è prevalentemente sensibile a sostanze dolci e a sostanze
salate, i lati della lingua a sostanze acide e la parte posteriore a sostanze
amare. Questa compartimentazione non è netta; infatti, ogni zona della lingua
è sensibile a tutte le qualità gustative, ma con diversa intensità della
risposta.
I microvilli dei recettori gustativi sono la sede della trasduzione degli
stimoli. Nel poro gustativo, le sostanze disciolte nella saliva entrano in
contatto con la porzione apicale della membrana delle cellule gustative. È da
notare che alcune sostanze a basso peso molecolare sono in grado di indurre
stimolazione gustativa anche dopo iniezione nel torrente circolatorio,
probabilmente per diffusione fino alla superficie apicale dei recettori. La
trasduzione avviene con modalità diverse a seconda della natura chimica della
sostanza. Nella maggior parte dei casi, ne consegue la depolarizzazione della
membrana (potenziale generatore) e l'innesco di potenziali d'azione nella
cellula gustativa primaria. Nella porzione basale del recettore gustativo,
l'aumento della concentrazione di Ca++ indotto dall'apertura di canali
voltaggio-dipendenti o dalla mobilizzazione di riserve intracellulari provoca il
rilascio di mediatori chimici e la conseguente trasmissione del segnale alle
fibre afferenti gustative primarie.
L'attività dei recettori gustativi può essere modulata ad opera di circuiti
locali e da fattori endocrini. Nei bottoni gustativi sono state messe in luce
giunzioni comunicanti (gap junctions) e sinapsi chimiche tra i diversi tipi
cellulari. Anche se la loro funzione non èinteramente nota, è ipotizzabile che
esse, consentendo un flusso laterale di informazioni, contribuiscano
all'elaborazione del segnale gustativo.
Vie afferenti e circuiti centrali.
Le fibre
afferenti che convogliano le inforinazioni gustative alle strutture centrali
appartengono a tre paia di nervi cranici (VII, IX, X) e possono avere funzioni
almeno in parte diverse a seconda della loro provenienza. Ogni fibra risponde in
genere a 2, 3 o tutte e 4 le qualità gustative, anche se con intensità
diversa.
La porzione anteriore della lingua è innervata da un ramo (corda del timpano)
del nervo facciale (VII). Un altro ramo sensoriale del facciale, il grande
petroso superficiale, innerva i bottoni gustativi della mucosa del palato. I
corpi cellulari di questi neuroni si trovano nel ganglio genicolato. I neuroni
gustativi del nervo facciale rispondono in modo particolare a sostanze
nutritive, e le informazioni da essi trasmesse possono contribuire ad innescare
l'ingestione del cibo. La porzione posteriore della lingua, gli archi
palatoglossi e parte dell'orofaringe sono innervati dal ramo linguale del nervo
glossofaringeo (IX), i cui neuroni si trovano nel ganglio petroso. I neuroni
gustativi del IX nervo cranico sono molto sensibili a sostanze tossiche e
possono mediare riflessi avversivi. L'estremità posteriore della lingua,
l'epiglottide, la laringe e la porzione superiore dell'esofago sono innervate
dalla branca laringea superiore del nervo vago (X), i cui neuroni si trovano nel
ganglio nodoso: essi sono sensibili agli acidi e all'acqua e costituiscono
verosimilmente la branca afferente del riflesso della deglutizione e di altri
riflessi che proteggono le vie aeree dall'aspirazione di cibo o di liquidi.
Occorre inoltre ricordare che fibre trigeminali (V) innervano la maggior parte
dei bottoni gustativi, oltre all'epitelio non specializzato dell'orofaringe.
Anche se la loro funzione è in gran parte sconosciuta, queste fibre
contribuiscono verosimilmente all' apprezzamento della temperatura e della
consistenza del cibo.
I rami centripeti delle fibre gustative primarie terminano a livello del bulbo
nella parte rostrale del nucleo del tratto solitario, dove ha sede il secondo
neurone della via gustativa (questa porzione del nucleo del tratto solitario
viene anche chiamata nucleo gustativo). La maggior parte delle fibre gustative a
partenza dal nucleo del tratto solitario raggiunge la porzione parvicellulare
del nucleo ventrale posteromediale del talamo (VPMpc), attraverso il tratto
teginentale centrale. Il nucleo ventrale è il relais talamico delle
sensibilità facciale e orale. Dal VPMpc le informazioni gustative sono
trasmesse all'insula anteriore, alla parte interna dell'opercolo frontale e ad
un'area di corteccia granulare situata nella porzione inferiore del giro
precentrale, sulla convessità laterale degli emisferi. Quest'ultima regione
costituisce parte dell'area 3b ed è contigua caudalmente con la corteccia
gustativa primaria; tuttavia, i neuroni che rispondono a stimoli gustativi sono
relativamente pochi (2-11 per cento dell'intera popolazione), e frammisti ad
altri attivati ad esempio dai movimenti o dalla stimolazione tattile della
lingua. La maggior parte delle cellule gustative dell'insula e dell'opercolo
frontale risponde a più di una qualità gustativa. Neuroni sensibili a stimoli
gustativi sono presenti nei primati anche nella porzione latero-posteriore della
corteccia orbitofrontale (aree 12 e 13 di Brodmann), che riceve proiezioni dall'insula
anteriore. I neuroni orbitofrontali hanno caratteristiche diverse e più
complesse: sono in genere attivati da una sola qualità gustativa, ma possono
rispondere anche a stimoli correlati in altre modalità sensoriali (ad esempio,
il sapore dolce e l'odore di frutta). Inoltre, nei neuroni sensibili a sostanze
nutritive, come il glucosio, la risposta viene inibita in maniera specifica
quando la scimmia ne è sazia. Queste caratteristiche suggeriscono un
coinvolgimento di quest'area nell'integrazione delle informazioni sensoriali e
nella regolazione del comportamento alimentare.
Nei primati, le vie gustative sono ipsilaterali fino a livello talamico. A
livello corticale, la stimolazione gustativa determina potenziali evocati
bilateralmente, anche se le risposte ipsilaterali sono più intense ed hanno
latenza minore. Nell'uomo, sono stati descritti deficit gustativi di un lato in
seguito a lesioni corticali sia ipsi- che contro-laterali.
Dal nucleo del tratto solitario partono anche fibre dirette ai nuclei salivatori
inferiore e superiore, che regolano l'attività delle ghiandole salivari
(riflesso di salivazione), e al complesso parabrachiale pontino, alcune porzioni
del quale rispondono a stimoli gustativi. Il complesso parabrachiale invia fibre
al nucleo centrale dell'amigdala, al nucleo interstiziale della stria terminale
e all'ipotalamo, oltre che alla sostanza reticolare del tronco dell'encefalo. I
circuiti a partenza dal complesso parabrachiale pontino sono probabilmente
implicati nelle risposte riflesse a stimoli gustativi quali la sudorazione, la
vasocostrizione e il riflesso di retrazione al gusto amaro. Inoltre, le
connessioni con l'ipotalamo potrebbero essere coinvolte nel convogliamento
dell'informazione gustativa ai centri della fame, della sazietà e della sete.
Le informazioni gustative iniluenzano il tono affettivo e l'attività dei
complessi meccanismi comportamentali, endocrini e vegetativi implicati nel
controllo della composizione chimica del "mezzo interno". Oltre alle
sopracitate proiezioni dal complesso parabrachiale, le connessioni dell'insula e
della corteccia orbitofrontale con altre strutture facenti parte del sistema
limbico, come la porzione anteriore della corteccia cingolata e l?amigdala, sono
verosimilmente importanti a questo riguardo.
Rappresentazione centrale degli stimoli gustativi.
Recenti osservazioni sull'esistenza di popolazioni di neuroni che rispondono
prevalentemente a determinati stimoli gustativi, e sul mantenimento di un certo
grado di suddivisione topografica attraverso il nucleo del tratto solitario, il
complesso parabrachiale pontino, la porzione parvicellulare del nucleo ventrale
posteromediale, fino all'area 3b, suggeriscono che la codifica e la
rappresentazione centrale della qualità gustativa avvengano, almeno
parzialmente, tramite l'attivazione di linee specifiche di trasmissione. Secondo
un'altra teoria, avvalorata dal fatto che la maggioranza dei neuroni centrali è
sensibile, anche se in misura diversa l'una dall'altra, a più sostanze dal
gusto diverso, l'identificazione della qualità gustativa sarebbe basata
sull'analisi del profilo globale di attivazione di una vasta popolazione di
neuroni. E probabile che entrambe le teorie siano valide: il riconoscimento di
specifici sapori deriverebbe dalla comparazione tra il grado di attivazione di
determinate linee e il pattern" globale di attività del sistema. La
codifica dell'intensità dello stimolo è plausibilmente legata alla frequenza
dei potenziali d'azione dei neuroni gustativi.
Stimoli gustativi e regolazione dell'assunzione di cibo
e dei processi digestivi
La stimolazione gustativa della mucosa orofaringea è in grado di attivare
diversi meccanismi coinvolti, oltre che nella determinazione della qualità e
dell'intensità dello stimolo e nell'innesco di riflessi somatici (deglutizione
o espulsione del bolo) o viscerali (salivazione, aumento della secrezione
gastrica), nella genesi della sensazione di piacere che accompagna il pasto e,
in parte, dei segnali che ne determinano l'inizio e la fine.
lì sapore influenza in modo determinante il grado di accettazione di una
sostanza, che è direttamente proporzionale alla piacevolezza della sensazione
gustativa. Ad esempio, le sostanze dolci, fonte di energia per l'organismo,
danno di solito una sensazione piacevole, mentre il gusto amaro, tipico della
maggior parte dei veleni organici, provoca una sensazione sgradevole e il
rifiuto della sostanza; se lo stimolo è molto intenso, può innescare il
riflesso del vomito.
Il grado di piacevolezza e l'assunzione di un cibo sono influenzati dalle
necessità nutrizionali del momento: lo stesso cibo può essere avvertito come
molto gradevole e desiderabile all'inizio del pasto e sgradevole quando viene
raggiunta la sazietà. Ricerche condotte sulle scimmie hanno messo in luce che
alcuni neuroni dell'ipotalamo e della corteccia orbitofrontale diminuiscono
progressivamente la loro attività durante il pasto. Esiste quindi un
adattamento centrale di alcuni circuiti gustati vi in relazione al grado di
sazietà e verosimilmente indotto da fattori gastrointestinali ed ematici che si
modificano nel corso del pasto. Recenti studi condotti su soggetti umani
dimostrano che l'assunzione di cibo può essere modificata anche dalla semplice
applicazione di stimoli gustativi sulla mucosa orale. Ad esempio, la
stimolazione con glucosio o maltosio riduce significativamente il consumo di
cereali.
Sperimentalmente, è possibile notare che gli animali mantenuti ad una dieta
priva di uno specifico nutriente hanno la capacità di scegliere il cibo più
ricco di tale componente. Questo fenomeno è chiamato fame specifica. Un altro
fenomeno che conferma l'importanza della sensibilità gustativa nell'assunzione
di determinate sostanze è la fame per i/ sale (NaCl). La maggior parte dei
mammiferi necessita di un apporto costante di sale tramite la dieta e, di norma,
l'assunzione di sale è in eccesso rispetto alle esigenze dell'organismo.
L'equilibrio idrosalino è mantenuto dall'azione di alcuni ormoni, tra i quali
ha grande importanza l'aldosterone, liberato dalla corteccia cortico-surrenalica
in condizioni fisiologiche e fisiopatologiche (come l'ipovolemia). È stato
dimostrato che l'aldosterone aumenta il trasporto di Na+ nella membrana delle
cellule gustative (come in altre cellule epiteliali, ad esempio a livello
renale), e la risposta al cloruro di sodio nelle fibre afferenti gustative della
corda del timpano. L'eccesso di stimolo salato tende così a ridurre
l'introduzione alimentare di sale. Infatti, in ratti surrenectomizzati, si può
notare una riduzione della sensibilità dei recettori a sostanze dal gusto
salato, e una netta preferenza compensatoria per i cibi ricchi di sale. Lo
stesso fenomeno è riscontrabile in pazienti con deficienze surrenaliche.
Le informazioni gustative sono biologicamente importanti anche per il controllo
dei processi digestivi. Intatti, la secrezione di molte ghiandole (salivari,
pancreatiche, gastriche, eccetera) viene attivata e modulata in via riflessa
attraverso le connessioni tra centri gustativi (nucleo del tratto solitario,
pars medialis del nucleo parabrachiale, porzione parvicellulare del talamo
ventroposteromediale) e nuclei implicati nel controllo delle efferenze
vegetative (sopratutto vagali). È importante notare che gli stimoli gustativi
influenzano sia la quantità che la composizione del secreto. Ad esempio,
stimoli acidi fanno aumentare la componente acquosa (e di HCO3) delle ghiandole
salivari e delle ghiandole di von Ebner: stimoli amari incrementano la
secrezione enzimatica delle ghiandole gastriche; la presenza di grassi nel cavo
orale stimola la secrezione delle lipasi salivari, gastriche e pancreatiche.
Infine, stimoli dolci sembrano influenzare la secrezione di ormoni ed enzimi
coinvolti nel metabolismo dei glucidi.
Psicobisiologia della sensibilità gustativa
Il senso del gusto consente di riconoscere parecchie centinaia di sapori
diversi. Come ricordato in precedenza, si ritiene che tutti i sapori percepibili
risultino dalla combinazione di quattro qualità gustative primarie (dolce,
amaro, salato, acido): secondo alcuni autori, il sapore del glutammato
monosodico, sostanza largamente impiegata come additivo alimentare, costituisce
una quinta qualità gustativa a se stante.
La maggior parte degli stimoli gustativi è costituita da molecole idrofile, non
volatili, efficaci ad alte concentrazioni nei mammiferi (da mM a M), che
includono sali, acidi, zuccheri, aminoacidi, e proteine. Le grosse molecole
lipidiche sono scisse dalla lipasi salivare in molecole di minor peso
molecolare, capaci di stimolare i recettori gustativi (i grassi sono importanti
per la sapidità del cibo). Sia il sapore dolce che il sapore amaro sono evocati
da una classe eterogenea di sostanze, la maggior parte delle quali di natura
organica, delle quali i prototipi sono rappresentati dal saccarosio (dolce) e
dal chinino (amaro). Il sapore salato è provocato da sali ionizzati, come il
cloruro di sodio. Il sapore acido è in relazione al pH della soluzione: tanto
minore è questo tanto più intensa è la sensazione. Per soglia gustativa di
una sostanza si intende la concentrazione minima in soluzione che determina una
sensazione gustativa. Come per le soglie olfattive, è necessario distinguere
tra soglia protopatica e soglia epicritica. La concentrazione sufficiente a
stimolare i recettori gustativi e ad indurre una sensazione gustativa (soglia
protopatica) è inferiore alla concentrazione alla quale il sapore viene
riconosciuto con sicurezza (soglia epicritica). È interessante ricordare che il
sapore può essere in relazione alla concentrazione. Ad esempio, il sale da
cucina a bassa concentrazione viene percepito come dolce, ad alta concentrazione
come amarognolo. La soglia gustativa è normalmente più bassa per il sapore
amaro, in accordo con quanto sopra ricordato sulla funzione di monitoraggio
della presenza di sostanze potenzialmente tossiche. Inoltre, le soglie gustative
presentano differenze legate al sesso. Le donne mostrano una maggiore
sensibilità rispetto agli uomini per gli stimoli dolci e salati, e una
sensibilità minore per gli stimoli acidi.
La sensibilità gustativa, così come quella olfattiva, è caratterizzata da un
rapido adattamento (cioè, una rapida diminuzione dell'intensità del sapore
percepito), che è già completo dopo un minuto dall'applicazione della molecola
stimolante. L'adattamento ad uno dei sapori fondamentali non ha effetti
significativi sulle risposte neurali e percettive agli altri sapori primari:
ciò conferma la presenza di meccanismi indipendenti di codifica ed elaborazione
delle diverse qualità gustative.
Deficit gustativi
Deficit generalizzati della funzione gustativa sono relativamente rari. Il senso
del gusto sembra più resistente di quello dell'olfatto agli effetti dell'età,
di malattie degenerative del sistema nervoso e di traumi cranici. Ciò può
essere legato a diversi fattori, sia periferici (le informazioni gustative sono
trasmesse da diversi nervi cranici, e in caso di lesione unilaterale le
strutture del lato sano sono sufficienti a prevenire l'insorgenza di deficit)
che centrali. I più importanti fattori clinici in grado di indurre alterazioni
generalizzate della funzione gustativa sono le condizioni che compromettono il
turnover dei recettori gustativi, quali infiammazioni (stomatiti) e lesioni del
cavo orale da agenti virali, da farmaci, o da radiazioni ionizzanti. Deficit
gustativi sono anche caratteristici del diabete. Alcune sostanze o farmaci
alterano in maniera specifica la percezione di stimoli salati (amiloride) o
dolci (acido gimnemico).
Si definisce ipogeusia la condizione nella quale la soglia di percezione del
gusto si innalza rispetto ai valori normali. Per ageusia si intende la perdita
totale della sensibilità gustativa.
Il termine disgeusia indica una sensibilità gustativa alterata (non
corrispondente allo stimolo), o la comparsa di sensazione gustativa in assenza
di stimoli. Fenomeni disgeusici possono manifestarsi in casi di neoplasie
dell'apparato digerente (in alcuni casi di tumori gastrici i pazienti rifiutano
la carne perché questa risulta di sapore sgradevole o amaro).
DEGLUTIZIONE
La deglutizione consiste in una
sequenza di contrazioni muscolari che trasportano il cibo dalla cavità orale
allo stomaco. Gli eventi preparatori della deglutizione (gli unici che possono
avere un qualche interesse stomatognatico) consistono nello spostare il bolo in
posizione centrale tra la parte anteriore della lingua ed il palato rigido. La
tappa seguente è quella della chiusura delle labbra (se non già avvenuta),
mentre la punta della lingua viene premuta verso il palato rigido. La mandibola
viene ancorata ed il palato molle elevato per occludere il rinofaringe ed aprire
le fauci, evitando così il reflusso degli alimenti all'interno del naso. Un
progressivo piegarsi a forma di arco verso l'indietro del corpo della lingua
contro il palato molle, sospinge il bolo verso la faringe.
  |
| Sequenza cinematografica di un
atto di deglutizione normale di un bolo semiliquido. |
Succesivamente, la parte posteriore
della lingua si sposta in avanti in modo tale da permettere al bolo di
penetrare nella faringe. A questo punto la respirazione viene momentaneamente
interrotta. La laringe si alza bruscamente e l'epiglottide si chiude in modo
tale da ostruire la via aerea. L'elevazione della laringe si accompagna ad uno
spostamento verso l'avanti dell'osso ioide. Contemporaneamente all'elevazione
dello ioide vi è un'azione di rilasciamento dello sfintere faringo-esofageo.
L'elevazione dello ioide è consentita dall'ancoraggio della mandibola da
parte dei suoi muscoli elevatori; la contrazione degli abbassatori inseriti
sullo ioide è quella che ne promuove l'innalzamento. I muscoli abbassatori
della mandibola ad inserzione iodea, quindi, sono attivi sia nella apertura
della mandibola che nella deglutizione. I diversi effetti meccanici sono
dovuti all'azione dei muscoli ad inserzione ioidea e sternale: se contratti,
l'osso ioide è fisso e la contrazione dei muscoli ioidei superiori promuove
abbassamento della mandibola (masticazione); se rilasciati, l'osso ioide è
mobile e la contrazione dei muscoli ioidei superiori ne promuove, a mandibola
fissa, l'innalzamento (deglutizione). I muscoli ioidei inferiori sarebbero
quindi da considerarsi muscoli masticatori, anche se non prendono diretto
rapporto con le parti mobili dell'apparato stomatognatico.
Durante la deglutizione si ha frequentemente un contatto interdentario in
intercuspidazione. Tale contatto ha una durata considerevolmente maggiore di
quella che intercorre alla fine dell'atto masticatorio: mediamente circa 680
msec per il primo e 195 per il secondo.
Normalmente, il bolo viene completamente ingurgitato nel corso di due o tre
movimenti deglutitori. L'intensità della contrazione dei muscoli masticatori
durante la deglutizione varia molto anche nello stesso soggetto e da un lato
all'altro: di norma il lato di masticazione preferenziale, dove la forza di
contrazione è maggiore durante la masticazione, è anche quello di maggiore
attività durante la deglutizione. Negli atti deglutitori più marcati sono
chiaramente distinguibili due fasi di contrazione del massetere. La durata
dell'atto deglutitorio varia da meno di un secondo a quasi due secondi.
Nel caso di un bolo solido, quindi, la deglutizione può essere suddivisa in
tre fasi:
prima fase: la lingua volge la punta in contatto con la mucosa palatina
dei denti anteriori e si apre a doccia fino a collabire con le pareti delle
arcate dentarie. I denti si distanziano un po' e le guance vengono contratte;
seconda fase: la lingua si contrae e si inarca dall'avanti
all'indietro, facendo scivolare posteriormente il bolo nel canale formato
dalla lingua stessa. In molti casi alla fine di questa fase si ha una leggera
separazione delle arcate dentarie, che consente una maggiore apertura della
cavità orale posteriormente e contribuisce all'instaurarsi di un leggero
gradiente pressorio (3-4 mm Hg) fra la parte anteriore e quella posteriore
della cavità orale;
terza fase: la laringe si chiude, impedendo il passaggio del contenuto
orale in trachea; segue un'ultima contrazione della lingua che spinge
violentemente il bolo in esofago.
Nell'atto del bere (come pure nella deglutizione della saliva) la successione
degli eventi é diversa. Si ha una maggiore azione delle guance che tendono a
creare una maggior negatività all'interno della cavità orale, mentre la
lingua non poggia con continuità sulla volta del palato, ma anzi forma un
canale. Il trasporto del contenuto avviene solo per negatività e per
gravità. Una più tardiva chiusura della laringe (dovuta forse ad una minore
stimolazione meccanica dei recettori dell'istmo delle fauci e responsabile,
forse, del più facile ingresso di liquidi che non di solidi in trachea)
consente un mantenimento più lungo della debole pressione negativa che
consente l'effetto di aspirazione sopra descritto. In alcuni casi, movimenti
della lingua di tipo simil-peristaltico possono facilitare il trasporto di
liquidi.
Nel neonato, la deglutizione si svolge con una meccanica ancora diversa, che
prende il nome di "comportamento deglutitorio di tipo infantile". Il
seno materno viene circondato dal labbro superiore e dalle gengive del
poppante superiormente, mentre inferiormente é la lingua che si distende
sopra alla gengiva inferiore e protrude tra capezzolo e labbro inferiore. La
spremitura del latte avviene grazie ad onde pressorie determinate da movimenti
ritmici della lingua, a sua volta sostenuta dalla mandibola.
RESPIRAZIONE
L'apparato stomatognatico non
partecipa normalmente alla respirazione, essendo l'ingresso d'elezione delle
vie aeree rappresentato dalle cavità nasali. Le attività fisiologiche
dell'apparato stomatognatico, però, influenzano vistosamente la meccanica
respiratoria, come nel caso della fonazione e, soprattutto, della masticazione
e, ancor più, della deglutizione. Durante la fonazione e la masticazione, il
ritmo respiratorio viene perlopiù rallentato e la espirazione prolungata. La
modificazione più vistosa, comunque, si ha durante la deglutizione, quando la
ventilazione viene interrotta da potenti riflessi che tendono a ridurre il
gradiente pressorio fra trachea ed istmo delle fauci, facilitando così
l'azione sigillante dell'epiglottide.
La respirazione orale è comune nella iperventilazione da sforzo (affanno), ma
ciò non crea normalmente problemi alla integrità o alla armonia funzionale
dell'apparato stomatognatico. Una respirazione orale abituale, una volta
instauratasi, può comportare danni quali crescita in post-rotazione della
mandibola per estrusione dei denti latero-posteriori, alterazioni della
postura linguale, iposviluppo del palato ed altro ancora.
FONAZIONE
Col termine di fonazione si intende,
nell'uomo, il complesso fenomeno di produzione della voce, derivante
dall'azione armonica degli organi fonatori, rappresentati dall'apparato
respiratorio, dall'apparato laringeo e dall'apparato risonatore o tubo di
risonanza, costituito dalle cavità nasale e orale, unica, quest'ultima, a
rivestire interesse di ordine stomatognatico. La voce, quale suono prodotto
dalle corde vocali, in concomitanza con rumori di tipo impulsivo o continuo
prodotti a vari livelli del tubo di risonanza o fonatorio, concorre alla
produzione dei fonemi: vocali e consonanti.
Le vocali si producono facendo passare la voce prodotta dalle corde vocali
attraverso la cavità di risonanza e l'articolazione buccale. Gli organi
articolatori principali sono rappresentati dalla lingua e dalle labbra.
Le consonanti sono il prodotto dell'aria sotto pressione proveniente dal
mantice polmonare che vince un ostacolo ostruente completamente il tubo
fonatorio (consonanti occlusive, come la T), oppure che passa attraverso la
fessura di un ostacolo che ostruisce solo parzialmente il tubo fonatorio
(consonanti costrittive o fricative, come la F), oppure ancora che vince prima
un ostacolo totalmente ostruente e poi uno parzialmente ostruente (consonanti
affricate, come la doppia C di caccia).
Gli ostacoli totali o parziali si creano di volta in volta per l'azione di un
organo articolatorio (labbro inferiore, lingua) che si mette in contatto o si
avvicina al contrapposto luogo di articolazione (labbro superiore, denti
superiori, palato duro, velo palatino). Se durante il fenomeno di occlusione
il velo pendulo é abbassato, parte dell'aria proveniente dal mantice
polmonare sfugge attraverso il tubo fonatorio secondario, rappresentato dalle
cavità nasali, e pertanto le occlusive corrispondenti si dicono nasali. In
caso contrario, le occlusive si dicono orali.
Qualsiasi evento che deterimini grossolane variazioni del volume del tubo
fonatorio o del suo orifizio esterno può determinare, se non compensato,
alterazioni della fonazione.
Da un punto di vista semeiologico, hanno una qualche importanza odontoiatrica
le lettere F e S. Durante la modulazione della F, infatti, il labbro inferiore
sfiora il margine incisale dei frontali superiori; tale fonazione può essere
utile per determinare soggettivamente l'altezza fisiologica degli elementi
dentali. Allo stesso modo, si può definire eccessiva l'altezza di elementi
dentari se essi entrano in contatto durante la fonazione della S (la distanza
normale a livello dei premolari é di circa 1-3 mm e corrisponde alla
posizione di riposo della mandibola).
GUSTO
La sensibilità gustativa
fornisce informazioni su alcune caratteristiche chimiche delle sostanze che
vengono a contatto con le mucose dell' orofaringe. I recettori gustativi
(cellule gustative primarie) interagiscono con le molecole disciolte nella
saliva e trasducono lo stimolo chimico in segnali nervosi. Il sapore è una
caratteristica dipendente dalla composizione della sostanza. Si ritiene che le
diverse centinaia di sapori percepibili derivino dalla combinazione di alcune
qualità primarie: dolce, salato, amaro e acido. I sapori primari differiscono
per la natura chimica dei composti in grado di evocarli e per i meccanismi di
captazione e di trasduzione. Contemporaneamente alla stimolazione gustativa,
molecole volatili liberate dalla sostanza possono legarsi ai recettori della
mucosa olfattiva. La sensazione globale che ne deriva è chiamata aroma.
L'importanza della sensibilità gustativa è collegata soprattutto alla
nutrizione e all'assunzione di liquidi. Il sapore di un cibo o di una bevanda
è infatti fondamentale nel determinarne la commestibilità e l'appetibilità;
il rifiuto di un cibo a causa del suo sapore sgradevole è importante per la
sopravvivenza di molte specie animali. Schematicamente, si possono associare
ai 4 sapori fondamentali funzioni diverse: assicurare le riserve energetiche
(dolce), mantenere l'equilibrio elettrolitico (salato), monitorare il pH
(acido, amaro), evitare sostanze tossiche (amaro). Di norma, le sostanze dolci
provocano una sensazione piacevole e l'innesco di riflessi di salivazione, di
deglutizione e di preparazione del tubo digerente alla digestione e
all'assorbimento. Invece, il sapore amaro provoca sovente il rifiuto del cibo
o della bevanda e, se molto intenso, stimola il riflesso del vomito: questo è
presumibilmente in relazione al fatto che la maggior parte delle sostanze
tossiche presenti in natura ha sapore amaro.
Il grado di piacevolezza di un sapore è tuttavia soggettivo e può essere
influenzato dall'esperienza e dalle necessità nutrizionali. Sia l'animale che
l'uomo tendono a rifiutare un cibo dal quale in passato siano stati
intossicati. In base alle informazioni gustative e olfattive si ha la
capacità di operare una scelta tra diversi cibi e preferire quello che
fornisce l'apporto nutritivo più consono alle esigenze dell'organismo.
Meccanismi e vie gustative
Struttura dell'apparato recettoriale.
I recettori gustativi sono cellule epiteliali specializzate, organizzate in
strutture dette bottoni gustativi.
I bottoni gustativi dei mammiferi
contengono alcune decine di recettori, oltre ad elementi cellulari immaturi
(cellule basali) La porzione apicale, dotata di microvilli, delle cellule
gustative primarie entra in contatto con la saliva che bagna la superficie
epiteliale mediante un canalicolo (poro gustativo). Tramite la porzione
basale, i recettori stabiliscono contatti sinaptici di tipo chimico con le
terminazioni di fibre afferenti primarie. Ogni fibra innerva numerosi bottoni
anche in zone dell'epitelio linguale molto distanti tra loro e, nello stesso
bottone, diversi recettori. Allo stesso tempo, ogni recettore gustativo è
innervato da più fibre afferenti. Morfologicamente, si possono distinguere
diversi tipi di recettori gustativi, cui corrispondono verosimilmente
differenze di tipo funzionale, ancora non chiarite. Le cellule basali
costituiscono una popolazione di cellule staminali che garantisce il ricambio
dei recettori gustativi (il ciclo vitale delle cellule gustative mature è di
circa dieci giorni, nei mammiferi).
I bottoni gustativi sono largamente diffusi nell'epitelio della lingua,
particolarmente nella sua porzione dorsale, ma sono presenti anche nella
mucosa del palato, della faringe, dell'epiglottide e del terzo superiore
dell'esofago. Nella lingua, sono per lo più localizzati nelle papille,
strutture specializzate di cui si distinguono diversi tipi: nell'uomo, solo le
papille vallate, fogliate e fungiformi hanno bottoni gustativi. Ognuna delle
papille vallate e fogliate contiene migliaia di bottoni. Le papille vallate
sono concentrate nella parte posteriore del dorso della lingua, a livello
della 'V" linguale. Le papille fogliate sono maggiormente presenti sui
lati della lingua. Le papille fungiformi si trovano numerose sulla punta della
lingua e contengono pochi bottoni gustativi: da I a 12 per papilla.
|
|
| A) Tipi di papille linguali.
Le papille fungiformi, foliate e vallate contengono bottoni gustativi.
B) Distribuzione delle aree linguali di maggior sensibilità |
Trasduzione degli stimoli gustativi.
La superficie recettoriale non risponde in modo omogeneo ai sapori primari. La
punta della lingua è prevalentemente sensibile a sostanze dolci e a sostanze
salate, i lati della lingua a sostanze acide e la parte posteriore a sostanze
amare. Questa compartimentazione non è netta; infatti, ogni zona della lingua
è sensibile a tutte le qualità gustative, ma con diversa intensità della
risposta.
I microvilli dei recettori gustativi sono la sede della trasduzione degli
stimoli. Nel poro gustativo, le sostanze disciolte nella saliva entrano in
contatto con la porzione apicale della membrana delle cellule gustative. È da
notare che alcune sostanze a basso peso molecolare sono in grado di indurre
stimolazione gustativa anche dopo iniezione nel torrente circolatorio,
probabilmente per diffusione fino alla superficie apicale dei recettori. La
trasduzione avviene con modalità diverse a seconda della natura chimica della
sostanza. Nella maggior parte dei casi, ne consegue la depolarizzazione della
membrana (potenziale generatore) e l'innesco di potenziali d'azione nella
cellula gustativa primaria. Nella porzione basale del recettore gustativo,
l'aumento della concentrazione di Ca++ indotto dall'apertura di canali
voltaggio-dipendenti o dalla mobilizzazione di riserve intracellulari provoca
il rilascio di mediatori chimici e la conseguente trasmissione del segnale
alle fibre afferenti gustative primarie.
L'attività dei recettori gustativi può essere modulata ad opera di circuiti
locali e da fattori endocrini. Nei bottoni gustativi sono state messe in luce
giunzioni comunicanti (gap junctions) e sinapsi chimiche tra i diversi tipi
cellulari. Anche se la loro funzione non èinteramente nota, è ipotizzabile
che esse, consentendo un flusso laterale di informazioni, contribuiscano
all'elaborazione del segnale gustativo.
Vie afferenti e circuiti centrali.
Le
fibre afferenti che convogliano le inforinazioni gustative alle strutture
centrali appartengono a tre paia di nervi cranici (VII, IX, X) e possono avere
funzioni almeno in parte diverse a seconda della loro provenienza. Ogni fibra
risponde in genere a 2, 3 o tutte e 4 le qualità gustative, anche se con
intensità diversa.
La porzione anteriore della lingua è innervata da un ramo (corda del timpano)
del nervo facciale (VII). Un altro ramo sensoriale del facciale, il grande
petroso superficiale, innerva i bottoni gustativi della mucosa del palato. I
corpi cellulari di questi neuroni si trovano nel ganglio genicolato. I neuroni
gustativi del nervo facciale rispondono in modo particolare a sostanze
nutritive, e le informazioni da essi trasmesse possono contribuire ad
innescare l'ingestione del cibo. La porzione posteriore della lingua, gli
archi palatoglossi e parte dell'orofaringe sono innervati dal ramo linguale
del nervo glossofaringeo (IX), i cui neuroni si trovano nel ganglio petroso. I
neuroni gustativi del IX nervo cranico sono molto sensibili a sostanze
tossiche e possono mediare riflessi avversivi. L'estremità posteriore della
lingua, l'epiglottide, la laringe e la porzione superiore dell'esofago sono
innervate dalla branca laringea superiore del nervo vago (X), i cui neuroni si
trovano nel ganglio nodoso: essi sono sensibili agli acidi e all'acqua e
costituiscono verosimilmente la branca afferente del riflesso della
deglutizione e di altri riflessi che proteggono le vie aeree dall'aspirazione
di cibo o di liquidi. Occorre inoltre ricordare che fibre trigeminali (V)
innervano la maggior parte dei bottoni gustativi, oltre all'epitelio non
specializzato dell'orofaringe. Anche se la loro funzione è in gran parte
sconosciuta, queste fibre contribuiscono verosimilmente all' apprezzamento
della temperatura e della consistenza del cibo.
I rami centripeti delle fibre gustative primarie terminano a livello del bulbo
nella parte rostrale del nucleo del tratto solitario, dove ha sede il secondo
neurone della via gustativa (questa porzione del nucleo del tratto solitario
viene anche chiamata nucleo gustativo). La maggior parte delle fibre gustative
a partenza dal nucleo del tratto solitario raggiunge la porzione
parvicellulare del nucleo ventrale posteromediale del talamo (VPMpc),
attraverso il tratto teginentale centrale. Il nucleo ventrale è il relais
talamico delle sensibilità facciale e orale. Dal VPMpc le informazioni
gustative sono trasmesse all'insula anteriore, alla parte interna
dell'opercolo frontale e ad un'area di corteccia granulare situata nella
porzione inferiore del giro precentrale, sulla convessità laterale degli
emisferi. Quest'ultima regione costituisce parte dell'area 3b ed è contigua
caudalmente con la corteccia gustativa primaria; tuttavia, i neuroni che
rispondono a stimoli gustativi sono relativamente pochi (2-11 per cento
dell'intera popolazione), e frammisti ad altri attivati ad esempio dai
movimenti o dalla stimolazione tattile della lingua. La maggior parte delle
cellule gustative dell'insula e dell'opercolo frontale risponde a più di una
qualità gustativa. Neuroni sensibili a stimoli gustativi sono presenti nei
primati anche nella porzione latero-posteriore della corteccia orbitofrontale
(aree 12 e 13 di Brodmann), che riceve proiezioni dall'insula anteriore. I
neuroni orbitofrontali hanno caratteristiche diverse e più complesse: sono in
genere attivati da una sola qualità gustativa, ma possono rispondere anche a
stimoli correlati in altre modalità sensoriali (ad esempio, il sapore dolce e
l'odore di frutta). Inoltre, nei neuroni sensibili a sostanze nutritive, come
il glucosio, la risposta viene inibita in maniera specifica quando la scimmia
ne è sazia. Queste caratteristiche suggeriscono un coinvolgimento di
quest'area nell'integrazione delle informazioni sensoriali e nella regolazione
del comportamento alimentare.
Nei primati, le vie gustative sono ipsilaterali fino a livello talamico. A
livello corticale, la stimolazione gustativa determina potenziali evocati
bilateralmente, anche se le risposte ipsilaterali sono più intense ed hanno
latenza minore. Nell'uomo, sono stati descritti deficit gustativi di un lato
in seguito a lesioni corticali sia ipsi- che contro-laterali.
Dal nucleo del tratto solitario partono anche fibre dirette ai nuclei
salivatori inferiore e superiore, che regolano l'attività delle ghiandole
salivari (riflesso di salivazione), e al complesso parabrachiale pontino,
alcune porzioni del quale rispondono a stimoli gustativi. Il complesso
parabrachiale invia fibre al nucleo centrale dell'amigdala, al nucleo
interstiziale della stria terminale e all'ipotalamo, oltre che alla sostanza
reticolare del tronco dell'encefalo. I circuiti a partenza dal complesso
parabrachiale pontino sono probabilmente implicati nelle risposte riflesse a
stimoli gustativi quali la sudorazione, la vasocostrizione e il riflesso di
retrazione al gusto amaro. Inoltre, le connessioni con l'ipotalamo potrebbero
essere coinvolte nel convogliamento dell'informazione gustativa ai centri
della fame, della sazietà e della sete.
Le informazioni gustative iniluenzano il tono affettivo e l'attività dei
complessi meccanismi comportamentali, endocrini e vegetativi implicati nel
controllo della composizione chimica del "mezzo interno". Oltre alle
sopracitate proiezioni dal complesso parabrachiale, le connessioni dell'insula
e della corteccia orbitofrontale con altre strutture facenti parte del sistema
limbico, come la porzione anteriore della corteccia cingolata e
l'amigdala,
sono verosimilmente importanti a questo riguardo.
Rappresentazione centrale degli stimoli gustativi.
Recenti osservazioni sull'esistenza di popolazioni di neuroni che rispondono
prevalentemente a determinati stimoli gustativi, e sul mantenimento di un
certo grado di suddivisione topografica attraverso il nucleo del tratto
solitario, il complesso parabrachiale pontino, la porzione parvicellulare del
nucleo ventrale posteromediale, fino all'area 3b, suggeriscono che la codifica
e la rappresentazione centrale della qualità gustativa avvengano, almeno
parzialmente, tramite l'attivazione di linee specifiche di trasmissione.
Secondo un'altra teoria, avvalorata dal fatto che la maggioranza dei neuroni
centrali è sensibile, anche se in misura diversa l'una dall'altra, a più
sostanze dal gusto diverso, l'identificazione della qualità gustativa sarebbe
basata sull'analisi del profilo globale di attivazione di una vasta
popolazione di neuroni. E probabile che entrambe le teorie siano valide: il
riconoscimento di specifici sapori deriverebbe dalla comparazione tra il grado
di attivazione di determinate linee e il pattern" globale di attività
del sistema. La codifica dell'intensità dello stimolo è plausibilmente
legata alla frequenza dei potenziali d'azione dei neuroni gustativi.
Stimoli gustativi e regolazione dell'assunzione di
cibo e dei processi digestivi
La stimolazione gustativa della mucosa orofaringea è in grado di attivare
diversi meccanismi coinvolti, oltre che nella determinazione della qualità e
dell'intensità dello stimolo e nell'innesco di riflessi somatici
(deglutizione o espulsione del bolo) o viscerali (salivazione, aumento della
secrezione gastrica), nella genesi della sensazione di piacere che accompagna
il pasto e, in parte, dei segnali che ne determinano l'inizio e la fine.
lì sapore influenza in modo determinante il grado di accettazione di una
sostanza, che è direttamente proporzionale alla piacevolezza della sensazione
gustativa. Ad esempio, le sostanze dolci, fonte di energia per l'organismo,
danno di solito una sensazione piacevole, mentre il gusto amaro, tipico della
maggior parte dei veleni organici, provoca una sensazione sgradevole e il
rifiuto della sostanza; se lo stimolo è molto intenso, può innescare il
riflesso del vomito.
Il grado di piacevolezza e l'assunzione di un cibo sono influenzati dalle
necessità nutrizionali del momento: lo stesso cibo può essere avvertito come
molto gradevole e desiderabile all'inizio del pasto e sgradevole quando viene
raggiunta la sazietà. Ricerche condotte sulle scimmie hanno messo in luce che
alcuni neuroni dell'ipotalamo e della corteccia orbitofrontale diminuiscono
progressivamente la loro attività durante il pasto. Esiste quindi un
adattamento centrale di alcuni circuiti gustati vi in relazione al grado di
sazietà e verosimilmente indotto da fattori gastrointestinali ed ematici che
si modificano nel corso del pasto. Recenti studi condotti su soggetti umani
dimostrano che l'assunzione di cibo può essere modificata anche dalla
semplice applicazione di stimoli gustativi sulla mucosa orale. Ad esempio, la
stimolazione con glucosio o maltosio riduce significativamente il consumo di
cereali.
Sperimentalmente, è possibile notare che gli animali mantenuti ad una dieta
priva di uno specifico nutriente hanno la capacità di scegliere il cibo più
ricco di tale componente. Questo fenomeno è chiamato fame specifica. Un altro
fenomeno che conferma l'importanza della sensibilità gustativa
nell'assunzione di determinate sostanze è la fame per i/ sale (NaCl). La
maggior parte dei mammiferi necessita di un apporto costante di sale tramite
la dieta e, di norma, l'assunzione di sale è in eccesso rispetto alle
esigenze dell'organismo. L'equilibrio idrosalino è mantenuto dall'azione di
alcuni ormoni, tra i quali ha grande importanza l'aldosterone, liberato dalla
corteccia cortico-surrenalica in condizioni fisiologiche e fisiopatologiche
(come l'ipovolemia). È stato dimostrato che l'aldosterone aumenta il
trasporto di Na+ nella membrana delle cellule gustative (come in altre cellule
epiteliali, ad esempio a livello renale), e la risposta al cloruro di sodio
nelle fibre afferenti gustative della corda del timpano. L'eccesso di stimolo
salato tende così a ridurre l'introduzione alimentare di sale. Infatti, in
ratti surrenectomizzati, si può notare una riduzione della sensibilità dei
recettori a sostanze dal gusto salato, e una netta preferenza compensatoria
per i cibi ricchi di sale. Lo stesso fenomeno è riscontrabile in pazienti con
deficienze surrenaliche.
Le informazioni gustative sono biologicamente importanti anche per il
controllo dei processi digestivi. Intatti, la secrezione di molte ghiandole
(salivari, pancreatiche, gastriche, eccetera) viene attivata e modulata in via
riflessa attraverso le connessioni tra centri gustativi (nucleo del tratto
solitario, pars medialis del nucleo parabrachiale, porzione parvicellulare del
talamo ventroposteromediale) e nuclei implicati nel controllo delle efferenze
vegetative (sopratutto vagali). È importante notare che gli stimoli gustativi
influenzano sia la quantità che la composizione del secreto. Ad esempio,
stimoli acidi fanno aumentare la componente acquosa (e di HCO3) delle
ghiandole salivari e delle ghiandole di von Ebner: stimoli amari incrementano
la secrezione enzimatica delle ghiandole gastriche; la presenza di grassi nel
cavo orale stimola la secrezione delle lipasi salivari, gastriche e
pancreatiche. Infine, stimoli dolci sembrano influenzare la secrezione di
ormoni ed enzimi coinvolti nel metabolismo dei glucidi.
Psicobisiologia della sensibilità gustativa
Il senso del gusto consente di riconoscere parecchie centinaia di sapori
diversi. Come ricordato in precedenza, si ritiene che tutti i sapori
percepibili risultino dalla combinazione di quattro qualità gustative
primarie (dolce, amaro, salato, acido): secondo alcuni autori, il sapore del
glutammato monosodico, sostanza largamente impiegata come additivo alimentare,
costituisce una quinta qualità gustativa a se stante.
La maggior parte degli stimoli gustativi è costituita da molecole idrofile,
non volatili, efficaci ad alte concentrazioni nei mammiferi (da mM a M), che
includono sali, acidi, zuccheri, aminoacidi, e proteine. Le grosse molecole
lipidiche sono scisse dalla lipasi salivare in molecole di minor peso
molecolare, capaci di stimolare i recettori gustativi (i grassi sono
importanti per la sapidità del cibo). Sia il sapore dolce che il sapore amaro
sono evocati da una classe eterogenea di sostanze, la maggior parte delle
quali di natura organica, delle quali i prototipi sono rappresentati dal
saccarosio (dolce) e dal chinino (amaro). Il sapore salato è provocato da
sali ionizzati, come il cloruro di sodio. Il sapore acido è in relazione al
pH della soluzione: tanto minore è questo tanto più intensa è la
sensazione. Per soglia gustativa di una sostanza si intende la concentrazione
minima in soluzione che determina una sensazione gustativa. Come per le soglie
olfattive, è necessario distinguere tra soglia protopatica e soglia
epicritica. La concentrazione sufficiente a stimolare i recettori gustativi e
ad indurre una sensazione gustativa (soglia protopatica) è inferiore alla
concentrazione alla quale il sapore viene riconosciuto con sicurezza (soglia
epicritica). È interessante ricordare che il sapore può essere in relazione
alla concentrazione. Ad esempio, il sale da cucina a bassa concentrazione
viene percepito come dolce, ad alta concentrazione come amarognolo. La soglia
gustativa è normalmente più bassa per il sapore amaro, in accordo con quanto
sopra ricordato sulla funzione di monitoraggio della presenza di sostanze
potenzialmente tossiche. Inoltre, le soglie gustative presentano differenze
legate al sesso. Le donne mostrano una maggiore sensibilità rispetto agli
uomini per gli stimoli dolci e salati, e una sensibilità minore per gli
stimoli acidi.
La sensibilità gustativa, così come quella olfattiva, è caratterizzata da
un rapido adattamento (cioè, una rapida diminuzione dell'intensità del
sapore percepito), che è già completo dopo un minuto dall'applicazione della
molecola stimolante. L'adattamento ad uno dei sapori fondamentali non ha
effetti significativi sulle risposte neurali e percettive agli altri sapori
primari: ciò conferma la presenza di meccanismi indipendenti di codifica ed
elaborazione delle diverse qualità gustative.
Deficit gustativi
Deficit generalizzati della funzione gustativa sono relativamente rari.
Il senso del gusto sembra più resistente di quello dell'olfatto agli effetti
dell'età, di malattie degenerative del sistema nervoso e di traumi cranici.
Ciò può essere legato a diversi fattori, sia periferici (le informazioni
gustative sono trasmesse da diversi nervi cranici, e in caso di lesione
unilaterale le strutture del lato sano sono sufficienti a prevenire
l'insorgenza di deficit) che centrali. I più importanti fattori clinici in
grado di indurre alterazioni generalizzate della funzione gustativa sono le
condizioni che compromettono il turnover dei recettori gustativi, quali
infiammazioni (stomatiti) e lesioni del cavo orale da agenti virali, da
farmaci, o da radiazioni ionizzanti. Deficit gustativi sono anche
caratteristici del diabete. Alcune sostanze o farmaci alterano in maniera
specifica la percezione di stimoli salati (amiloride) o dolci (acido gimnemico).
Si definisce ipogeusia la condizione nella quale la soglia di percezione del
gusto si innalza rispetto ai valori normali. Per ageusia si intende la perdita
totale della sensibilità gustativa.
Il termine disgeusia indica una sensibilità gustativa alterata (non
corrispondente allo stimolo), o la comparsa di sensazione gustativa in assenza
di stimoli. Fenomeni disgeusici possono manifestarsi in casi di neoplasie
dell'apparato digerente (in alcuni casi di tumori gastrici i pazienti
rifiutano la carne perché questa risulta di sapore sgradevole o amaro).

|