Gli acidi nucleici
La struttura
Duplicazione
Trascrizione, trasporto e traduzione
Gli anni '50 segnarono anche tappe importanti nella ricerca biochimica per la scoperta del DNA.
Il DNA e l'RNA sono gli acidi nucleici. Essi rappresentano la terza
classe di biopolimeri fondamentali insieme a proteine e carboidrati.
Il DNA si trova nel nucleo delle cellule eucariote nel citoplasma delle
procariote (sottoforma di DNA circolare), mentre l'RNA si trova
abbondantemente nel citoplasma di tutti tipi di cellule. Isolati nel
1870, solo nel 1953 gli scienziati Watson e Crick riuscirono ad
elaborare un modello funzionale tridimensionale di entrambi.
La loro funzione è principalmente quella di depositari delle
informazioni genetiche, cioè della sintesi di tutte le
proteine
prodotte all'interno della cellula.
Il DNA è il vero e proprio depositario di tali informazioni,
costituito da migliaia di geni, sequenze di nucleotidi ognuno
codificante un determinato polipeptide.
Ci sono, invece, diversi tipi di RNA ognuno con una funzione specifica
ma tutti impegnati nella fase di duplicazione, trasporto e sintesi.
La parte elementare di un acido nucleico è il nucleotide:
esso
è costituito da un gruppo fosfato, uno zucchero e una delle
5
basi azotate.
Gli zuccheri costituenti gli acidi nucleici sono 2 aldopentosi: il
D-ribosio (negli RNA) e il 2-desossi-ribosio (nel DNA):

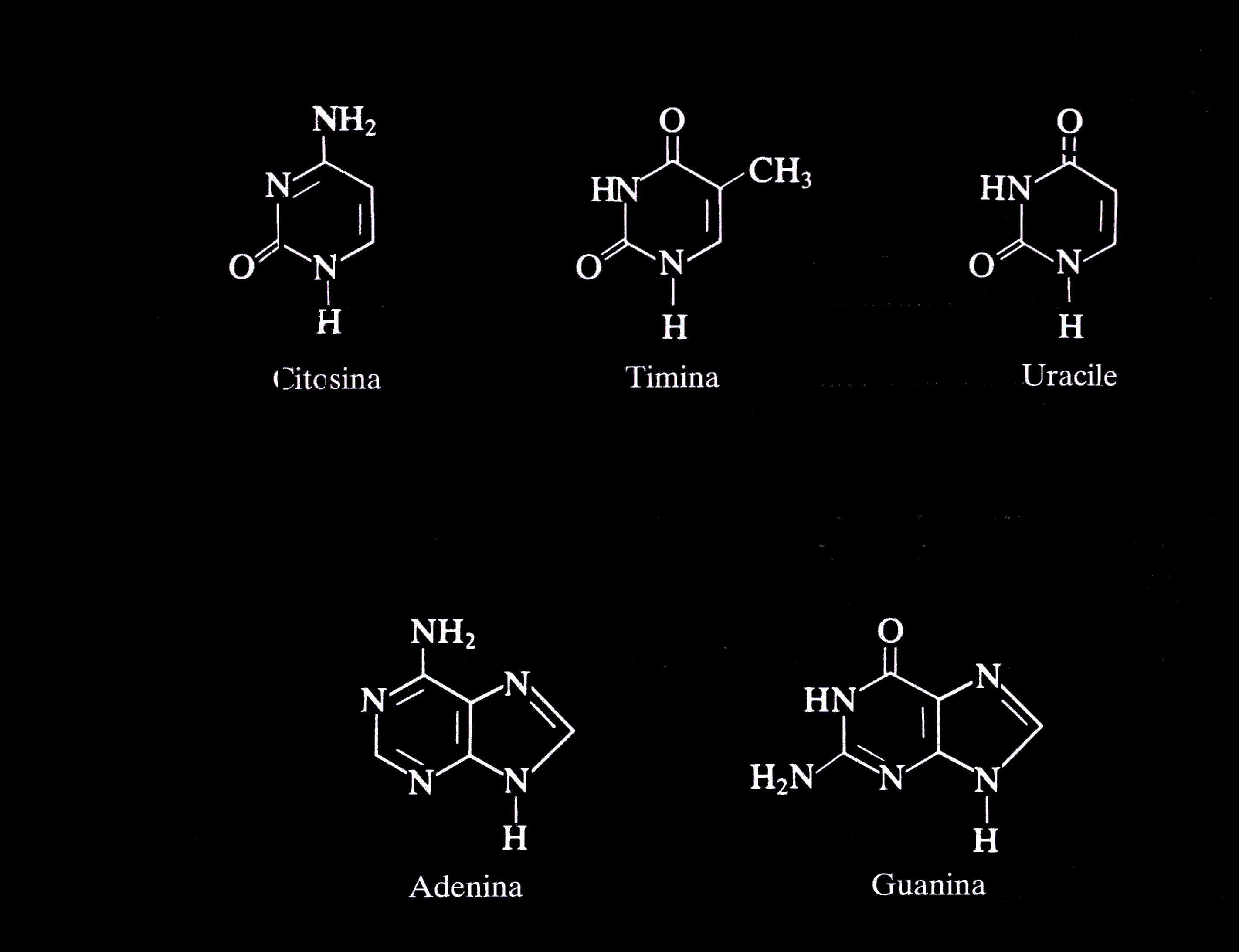
Le basi azotate, invece, derivano tutte da due basi eterocicliche fondamentali, la pirimidina e la purina:

Da esse, quindi derivano le varie
basi azotate. Dalla pirimidina derivano la citosina (DNA e RNA), la
timina (DNA) e l'uracile (RNA).
Dalla purina, invece, derivano l'adenina e la guanina, presenti
entrambi sia nell'RNA che nel DNA.
Gli zuccheri e
le basi azotate si uniscono mediante legame N-β- glicosidico
tra l'atomo di azoto (N9 nelle puriniche, N1 nelle pirimidiniche) e il
gruppo OH dello zucchero. La molecola risultante sarà un
nucleoside.

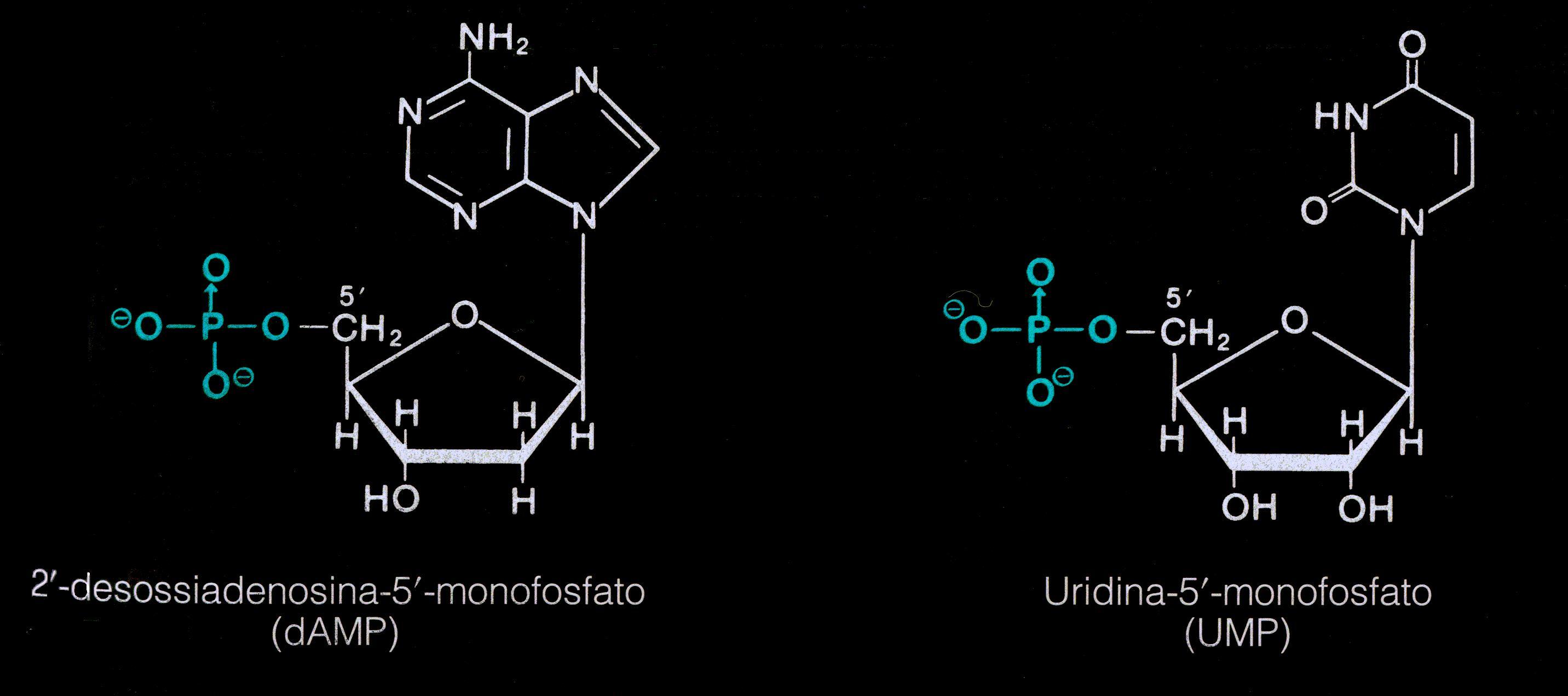
I
nucleotidi derivano da un'esterificazione del C5 dello zucchero con un
gruppo fosfato.
A seconda della natura dello zucchero essi si dividono in
desossiribonucleotidi e ribonucleotidi.
I
nucleotidi, come anche i nucleosidi, sono molto importanti anche in
forma singola. Si ricordano infatti i composti ATP (Adenosintrifosfato,
formato da tre gruppi fosfato legati ad un ribosio, a sua volta legato
all'adenosina) e l'ADP ( Adenosindifosfato, identico all'ATP ma con un
gruppo fosfato in meno) i quali sono le principali riserve di energia
vista la grande forza dei legami tra i legami tra i gruppi fosfato.
Per formare il DNA, migliaia e talvolta milioni di nucleotidi si legano
tra loro mediante legami fosfodiesterei che legano il doppietto di
elettroni sull'O del C3 all'O- del gruppo fosfato.
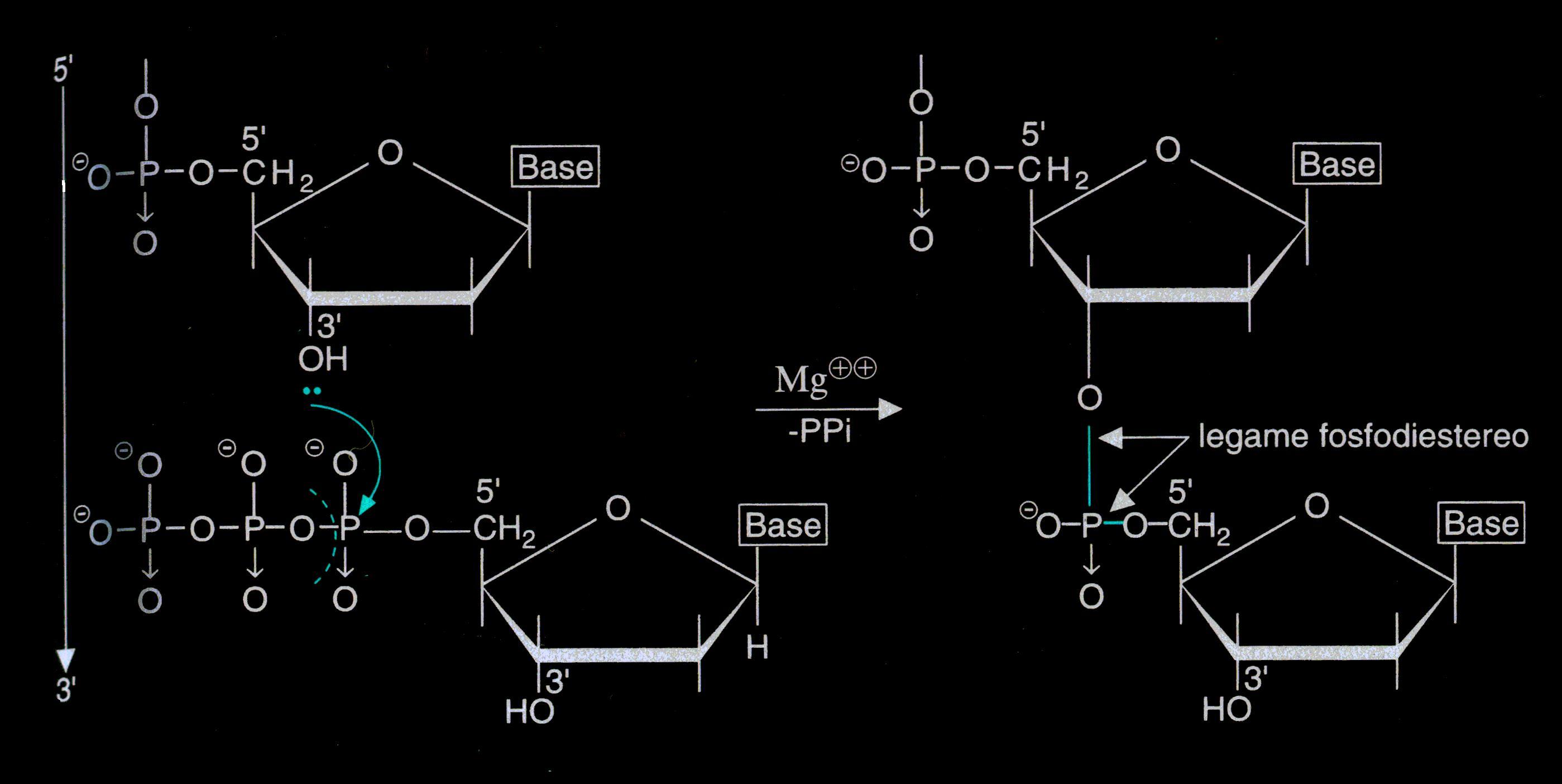
La
struttura primaria del DNA è una catena sequenziale di
nucleotidi legati da legami fosfodiesterei.
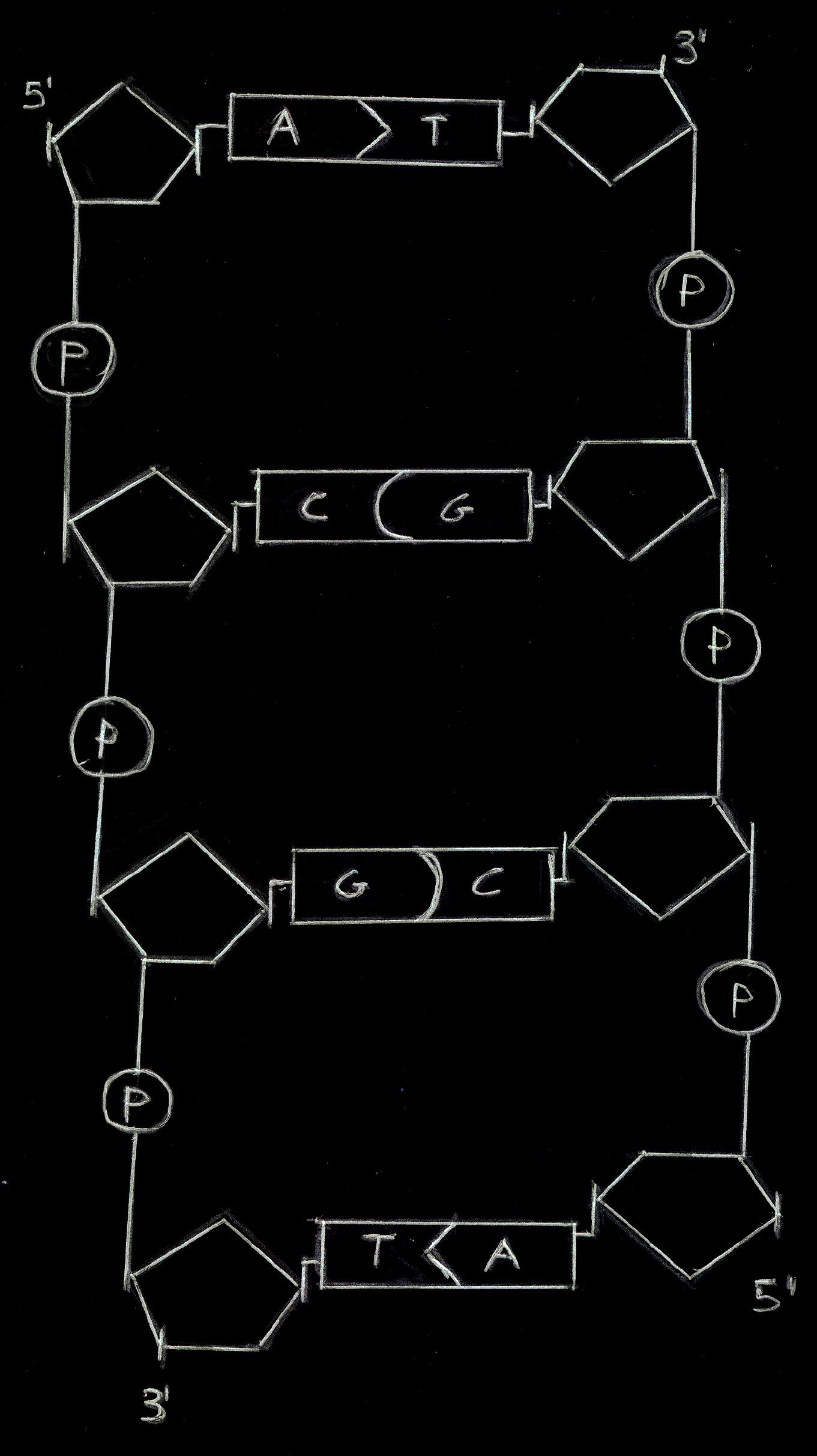
Il filamento viene detto Filamento 5'-3' se inizia col C5 per terminare
col C3 del desossiribosio. Viene detto Filamento 3'-5' se è
disposto al contrario.
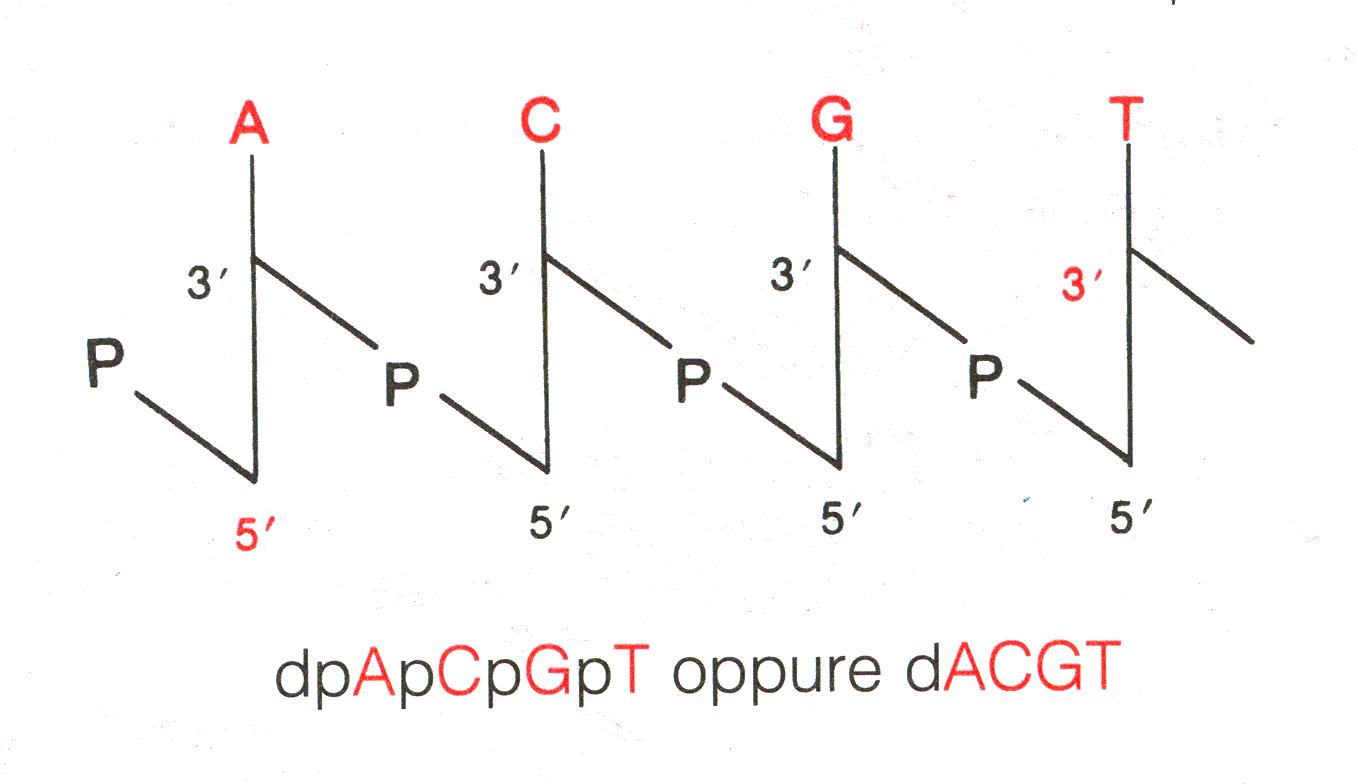
Nello
schema (un filamento 5'-3'), lo zucchero è una linea
verticale, il legame fosfodiestereo una verticale e le basi azotate
vengono indicate con le iniziali.
In natura, il DNA difficilmente si presenta sottoforma di singolo
filamento. Le sue successive organizzazioni sono tuttora in corso di
studio ma è stato dimostrato, con diffrazioni di raggi X,
che il DNA si presenta sottoforma di doppia elica tridimensionale,
esattamente come Watson e Crick teorizzarono.
I due filamenti complementari 5'-3' e 3'-5' si organizzano l'uno di
fronte all'altro, rispettando la complementarità e
ciò dà origine al doppio filamento che, per via
dei legami a idrogeno tra parti diverse della macromolecola, si ripiega
in una doppia elica.
La complementarità è dovuta a dei legami che
vengono a formarsi tra le basi azotate. In realtà,
più che legami sono dei ponti a idrogeno che si vengono a
formare tra le coppie A-T e C-G.
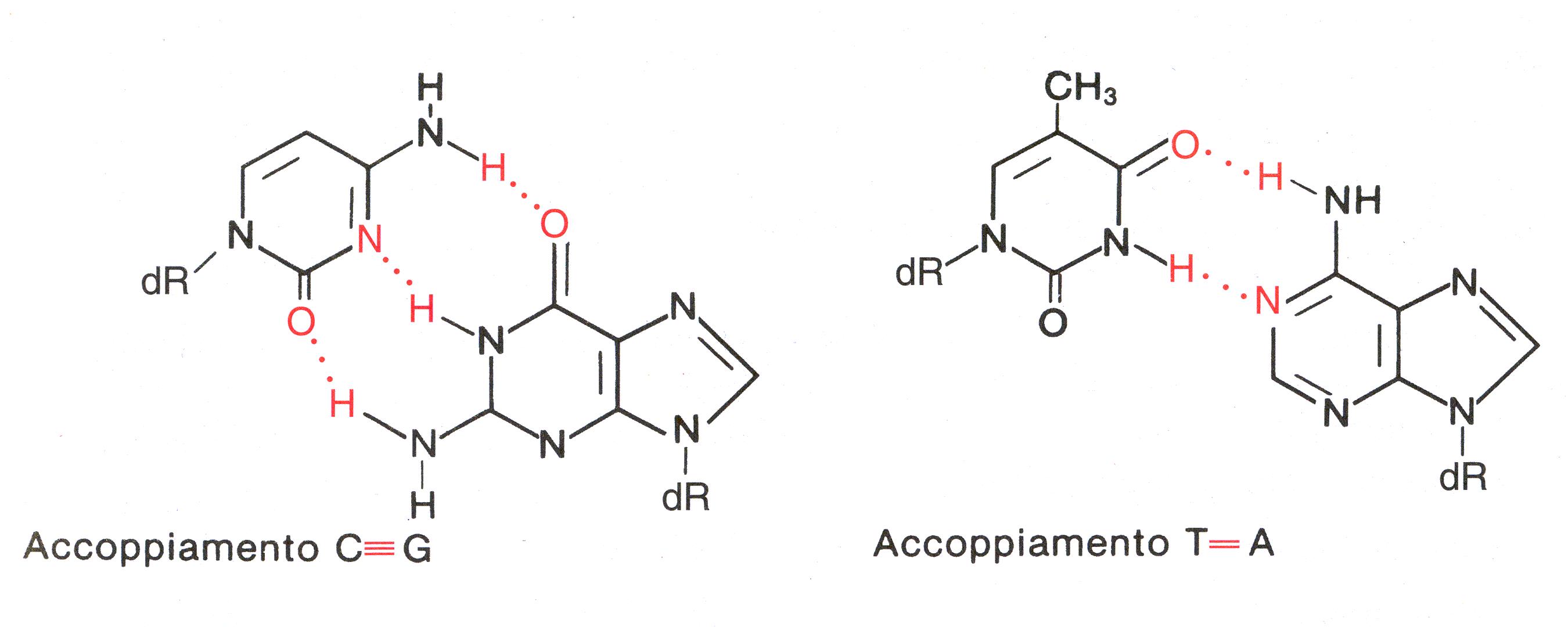
Il DNA si struttura quindi in una sequenza lineare, ripiegata da legami a idrogeno e complementare ad un altro filamento opposto.
Una cellula che sta per attraversare la mitosi ha bisogno di duplicare
il proprio filamento di DNA in modo da fornire alle due cellule figlie
un doppio filamento completo. Essendo i due filamenti complementari si
potrebbe intendere ogni filamento come stampo utilizzabile nella copia
dell'altro. Il risultato è una cellula con metà
DNA della cellula madre e l'altra metà sintetizzata al
momento.
La duplicazione di DNA è un processo di assemblaggio di
nucleotidi (una polimerizzazione) catalizzata da enzimi. L'enzima che
favorisce l'unione di nucleotidi è la DNA polimerasi.
Un enzima, l'elicasi, scioglie il doppio filamento, creando una
"bolla", contenente 2 forcelle di duplicazione in cui inizia la
duplicazione. Tali forcelle, col procedere dell'operazione, si
allargano sempre più fino a sciogliersi definitivamente
generando le due nuove molecole di DNA.
Nel filamento 5'-3', la reazione avviene normalmente e in maniera
continua. Nel filamento 3'-5' la DNA polimerasi può soltanto
unire nucleotidi ad un tratto già esistente, in quanto essa
non è in grado di aggiungere nucleotidi al filamento 5'.
Questo tratto viene chiamato primer. La DNA polimerasi aggiunge
nucleotidi ad uno stampo di primer già presente e completa
la duplicazione. Il primer è sintetizzato solo dopo che la
forcella di duplicazione ha lasciato liberi circa mille nucleotidi; la
reazione è catalizzata da un altro enzima, la primasi, che
duplica -senza bisogno,come la DNA polimerasi di un filamento
già presente- il filamento stampo, creando un piccolo tratto
di RNA , detto RNA primer, alle cui estremità 3' la DNA
polimerasi lega i nucleotidi. La presenza dell'RNA primer permette alla
DNA polimerasi di avviare il processo di duplicazione. Nei due
filamenti, quindi, la duplicazione avviene contemporaneamente ma con
direzione opposta.
In questo modo, sul filamento 3'-5', la reazione procede a piccoli
tratti e vengono sintetizzati dei frammenti di DNA (detti frammenti di
Okazaki) e non un filamento unitario come nel filamento 5'-3'. La DNA
polimerasi elimina i tratti di RNA primer e un altro enzima, la DNA
ligasi, provvede a legare tra loro i frammenti di Okazaki, ottenendo un
filamento continuo e complementare.
Trascrizione,
trasporto, traduzione
Se il DNA ha funzione di custodire il patrimonio genetico
dell'individuo, così l'RNA ha il compito di trasformare le
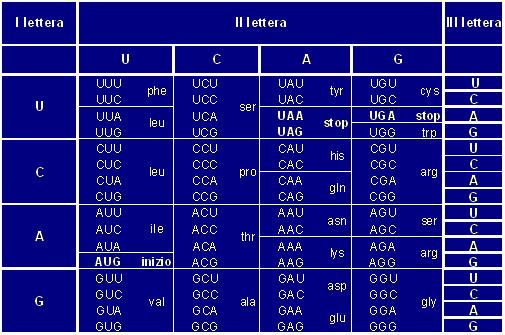
informazioni genetiche in azioni corrispondenti. Così una
serie di 3 nucleotidi (detta codone) sintetizza ognuna uno specifico
amminoacido. La trascrizione delle informazioni, dunque, parte dal DNA
che, mediante la trascrizione darà origine ad un mRNA in
grado di uscire dal nucleo cellulare ed essere letto dal ribosoma che,
decodificando il linguaggio genetico dei codoni, lo traduce in un
polipeptide.
L'espressione genica inizia nel momento in cui l'enzima RNA polimerasi catalizza la duplicazione di un filamento di DNA in uno di mRNA.
L'RNA polimerasi non necessita di un primer esistente come la DNA polimerasi e, inoltre, il processo avviene in una sola direzione. Esso parte da una sequenza di DNA, detta promotore, la quale viene riconosciuta dall'enzima RNA polimerasi e dà inizio al processo, che si conclude quando l'enzima incontra un'altra sequenza di DNA che, una volta riconosciuto, stoppa la trascrizione. Tale sequenza è detta terminatore.
Il filamento di mRNA risultante non è completo (infatti, viene detto pre-mRNA) in quanto contiene numerose sequenze che non codificano nessuna proteina. Tali sequenza sono dette introni e si distinguono dagli esoni, tratti codificanti proteine. Gli introni subiscono varie trasformazioni per essere rimossi e gli esoni vengono riassemblati per formare l'mRNA maturo. In realtà, più del 98% di un qualunque gene è formato da introni, la cui funzione è tuttora sconosciuta.
I ribosomi sono degli organuli presenti in qualunque tipo di cellula. Essi sono formati da una subunità minore ed una maggiore. Ogni subunità contiene rRNA (RNA ribosomiale) e proteine. L'rRNA è molto diverso dall'mRNA in quanto a lunghezza (fissa nel primo e variabile nel secondo) ed è molto simile in quasi tutte le specie. La funzione dei ribosomi è quella di catalizzare i legami peptidici tra gli amminoacidi che man mano vengono a formarsi, leggono le triplette di nucleotidi dell'mRNA e riconoscono i punti d'inizio e fine traduzione. Ci sono, infatti, dei codoni di inizio e fine che permettono di non creare confusione nella lettura; dopo il codone d'inizio, infatti, la fase di lettura procede di tre in tre.
Gli amminoacidi non si legano direttamente al codone. Infatti, esiste un terzo tipo di RNA (tRNA) il cui compito è quello di fungere da "appiglio" per il codone e per l'amminoacido.
Il tRNA è strutturato come una specie di trifoglio il cui fusto è costituito da basi azotate tenute assieme da legami a idrogeno. All'estremità del fusto vi è il sito di attacco per l'amminoacido mentre nella "foglia" opposta al fusto vi è una serie di tre basi azotate, complementari al codone destinato a quel tRNA. Queste tre basi azotate costituiscono l'anticodone.
Ogni tRNA è specializzato esclusivamente nel trasporto e nell'attacco di un particolare codone e, quindi, di un particolare amminoacido. Ogni cellula deve avere un minimo di venti tRNA diversi (una per ogni amminoacidi); il numero può variare sino a 61 (uno per ogni codone) dal momento che più codoni possono codificare un singolo amminoacido.
Se il codone si lega al tRNA per complementarità di basi azotate, l'amminoacido si lega ad esso grazie all'aiuto di particolari enzimi. Infatti, tali enzimi hanno un doppio punto d'attacco (siti). La struttura tridimensionale del tRNA permette all'enzima di riconoscere solo quel tRNA e di poter esporre solo all'amminoacido correlato solo il sito corrispondente e complementare.
La fase della traduzione si divide in tre momenti: l'inizio, l'allungamento, la terminazione:
1 Inizio: la traduzione inizia con l'aggancio del tRNA d'inizio alla subunità minore del ribosoma. L'unico codone che dà inizio alla traduzione è l'AUG, codificante la metonina. Ogni peptide appena creato inizia quindi con la metonina, che molte volte viene in seguito eliminata. L'mRNA viene steso e preparato per la traduzione.
2 Allungamento: questa fase prevede l'entrata in gioco della subunità maggiore del ribosoma. Quest'ultima ha due siti, il sito A e il sito P. il primo è destinato al tRNA nuovo che si lega al codone e che trasporta l'amminoacido, il secondo è destinato al peptide in formazione. L'allungamento procede per tre fasi: all'inizio, la metonina (o tutto il peptide nel sito A), si sposta sul sito P, lasciando libero il sito A su cui si aggancia il secondo tRNA con un altro amminoacido che, per l'intervento di alcuni enzimi, crea un legame peptidici con quello che lo precede. La seconda fase vede la partenza del primo tRNA (quello sul sito A); quel tRNA, però, stacca dal proprio sito l'amminoacido, lasciando tutto il peptide legato al secondo tRNA. La terza fase vede lo spostamento del tRNA con tutto il peptide in formazione dal sito A al sito P mentre il sito A viene occupato da un nuovo tRNA carico. Il processo si ripete finché non arriva un codone stop.
3 Terminazione: quando il ribosoma raggiunge un codone stop, la traduzione si ferma e sul sito A si lega una proteina, il fattore di rilascio, che permette ad alcuni enzimi di intervenire per staccare il peptide completo dal tRNA.
 |
 |
 |
 |