Meccanismi molecolari di trasduzione del segnale
I recettori sono proteine
di membrana altamente specifici che legano
selettivamente, in un sito 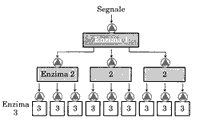 esterno, le molecole del segnale mediante
interazioni chimici deboli. La capacità dei recettori di
identificare e di legarsi alle molecole del segnale
è talmente elevata che queste interazioni possono avvenire
anche quando la concentrazione del ligando [segnale] è molto
piccola. Una volta legati, il complesso recettore-molecola del segnale
innesca un meccanismo di amplificazione dovuta all'azione di un enzima
legato al recettore che provoca una cascata enzimatica, attivando molte
molecole di un secondo enzima e poi di un terzo e così via.
[vedi figura]
esterno, le molecole del segnale mediante
interazioni chimici deboli. La capacità dei recettori di
identificare e di legarsi alle molecole del segnale
è talmente elevata che queste interazioni possono avvenire
anche quando la concentrazione del ligando [segnale] è molto
piccola. Una volta legati, il complesso recettore-molecola del segnale
innesca un meccanismo di amplificazione dovuta all'azione di un enzima
legato al recettore che provoca una cascata enzimatica, attivando molte
molecole di un secondo enzima e poi di un terzo e così via.
[vedi figura]
I sistemi di trasduzione possono essere integrati tra loro potendo ricevere segnali multipli e produrre un'unica risposta. Inoltre il prolungamento oltre le normali necessità della trasmissione del segnale si può interrompere perchè il recettore, dopo un po' di tempo si desensibilizza per un meccanismo a feedback in cui un enzima della cascata della risposta può produrre una inibizione retroattiva.
Le vie di trasduzione del segnale hanno caratteristiche comuni che possono essere così riassunte:
- un segnale interagisce con il suo recettore specifico
- il recettore attivato interagisce con un sistema molecolare interno che produce un nuovo segnale oppure la modificazione dell'attività di una proteina cellulare.
- si ha una modificazione dell'attività metabolica della cellula.
- la trasduzione
termina e la cellula ritorna nelle condizioni in cui era prima
dell'arrivo del segnale.
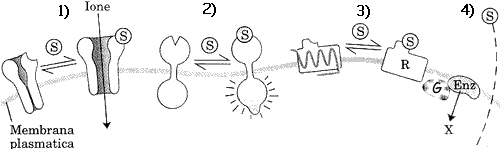
canali ionici: fig.1) sono canali presenti in proteine di membrana che
si possono aprire o chiudere in risposta ad una determinata molecola
del segnale (ligando).
A questa categoria appartengono anche le proteine recettrici fig.3) (non enzimatiche) che, indirettamente regolano, attraverso altre proteine (proteina G), legate al GTP, analogo dell'ATP, l'attività di enzimi che producono un secondo messaggero intracellulare. (X)
recettori nucleari: fig.4) essi si associano al loro ligando, come gli
ormoni steroidei (S), modificando la velocità di
trascrizione di un gene.
Canali ionici: i recettori dell'acetilcolina.
L'acetilcolina (Ach) è un neurotrasmettitore che viene liberato, da un neurone eccitato, nello spazio sinaptico e trova il suo recettore nella membrana postsinaptica di una cellula muscolare o di una nervosa. L'Ach propaga l'impulso elettrico (potenziale d'azione) proveniente dal corpo cellulare di un neurone. Quando la depolarizzazione della membrana (interno positivo) giunge alla terminazione di un assone, si aprono i canali proteici per il Ca++ la cui elevata concentrazione determina il rilascio di Ach nello spazio sinaptico. L'acetilcolina si lega ai recettori del neurone postinaptico causando l'apertura dei canali ionici per il Na+ e propagando il potenziale d'azione.
Recettori enzimatici: il recettore dell'insulina
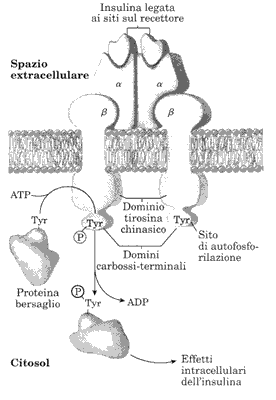 Il recettore dell'insulina, ormone pancreatico prodotto dalle ghiandole
β del pancreas, è una proteina complessa composta
da 4 subunità, due α collocate esternamente sulla
superficie della membrana e due β che attraversano la membrana
e sporgono nel citosol. [vedi figura]. Una catena α con la
catena β corrispondente formano un monomero.
Il recettore dell'insulina, ormone pancreatico prodotto dalle ghiandole
β del pancreas, è una proteina complessa composta
da 4 subunità, due α collocate esternamente sulla
superficie della membrana e due β che attraversano la membrana
e sporgono nel citosol. [vedi figura]. Una catena α con la
catena β corrispondente formano un monomero.
L'enzima ora è in grado di provocare la fosforilazione di alcuni residui di Tyr presenti in proteine bersaglio. Avviene così una cascata di fosforilazioni che coinvolge alcune proteine citosoliche che provocheranno gli effetti intracellulari dell'insulina come, ad esempio, aprire i canali per il trasporto del glucosio all'interno della cellula ed espletare l'effetto ipoglicemizzante dell'ormone.
Recettori che attivano un secondo messaggero intracellulare: il recettore β adrenergico dell'adrenalina
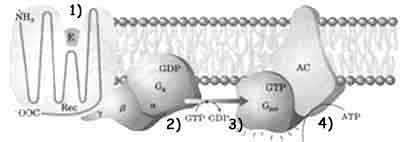 Il recettore per l'ormone adrenalina (β adrenergico)
è una proteina le cui 7 eliche attraversano la
membrana [vedi figura] formando quello che viene
chiamato il recettore a serpentina.
Il recettore per l'ormone adrenalina (β adrenergico)
è una proteina le cui 7 eliche attraversano la
membrana [vedi figura] formando quello che viene
chiamato il recettore a serpentina.
Il legame con l'adrenalina (1) modifica la conformazione del recettore che diviene in grado di catalizzare la sostituzione del GDP (2), legato ad una proteina stimolatrice, Gs, con il GTP. La proteina stimolatrice, composta da tre unità monomeriche, può così lasciare le subunità β e γ e, spostandosi lungo il piano della membrana con la sola subunità α, raggiungere una proteina integrale di membrana chiamata adenil ciclasi (3) attivandola. Il sito attivo della AC è rivolto verso il citosol e catalizza la conversione dell'ATP in AMPc (AMP ciclico).(vedi figura a sinistra dettaglio 4)
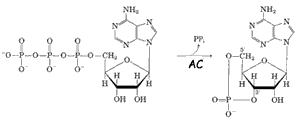 L'AMPc è un messaggero secondario che nel citosol stimola
l'azione di una proteina chinasi AMPc dipendente (PKA) in grado di
fosforilare le proteine cellulari e causare la risposta all'adrenalina.
L'AMPc è un messaggero secondario che nel citosol stimola
l'azione di una proteina chinasi AMPc dipendente (PKA) in grado di
fosforilare le proteine cellulari e causare la risposta all'adrenalina.