Introduzione: la comunicazione intercellulare
I numerosi processi cellulari di natura elementare, sono
regolati da
una vasta gamma di ormoni, neurotrasmettitori e sostanze paracrine.
La prima fase che consente il funzionamento di queste sostanze
è dato
dal loro legame con specifici recettori. I recettori sono di natura
proteica, posti nella membrana plasmatica di apposite cellule
bersaglio. Nella maggior parte dei casi, il recettore, è un
canale
ionico, modulato dal ligando (ormone, neurotrasmettitore ecc.) e, la
risposta della cellula, è una corrente ionica. Il canale
ionico diviene
dunque il recettore, ma anche l’effettore della risposta
cellulare.
Le sostanze regolatrici, che si legano ai recettori, sono classificate in 3 categorie:
sostanze endocrine (ormoni) sono liberate da cellule endocrine e sono in grado di raggiungere cellule bersaglio poste anche a notevole distanza rispetto al sito di secrezione, sfruttando il torrente ematico per essere veicolate;
sostanze neurocrine: vengono liberate da neuroni ed agiscono nelle immediate vicinanze del sito di secrezione. Appartengono a questa categoria i neurotrasmettitori e buona parte dei neuromodulatori;
sostanze paracrine: vengono liberate ad una distanza tale che possano raggiungere per diffusione le cellule bersaglio (es.: istamina).
Lo stadio finale delle vie di trasduzione del segnale
è quasi sempre
rappresentato dalla fosforilazione di una proteina avente una funzione
fondamentale ai fini di un dato processo biologico. La fosforilazione
delle proteine è compito delle proteine chinasi
cellulari, le proteine fosfatasi si occupano invece
di catalizzare il distacco del fosforo dalle proteine.
Tra i segnali che regolano l’attività delle
proteine chinasi ricordiamo: i secondi messaggeri AMPc,
GMPc, Ca++, inositolo trifosfato (IP3)
e alcuni di gliceridi.
All’interno delle cellue è possibile riscontrare
proteine chinasi
regolate da ognuno di questi messaggeri. Pertanto il legame della
molecola regolatrice al recettore determina variazioni dei livelli
intracellulari di uno dei secondi messaggeri. Alcune cellule possiedono
proteine chinasi la cui attività è aumentata
dall’AMPc e dal GMPc,
altre possiedono proteine chinansi aumentano la loro
attività se legate al complesso Ca++-calmodulina
(la calmodulina è presente in tutte le cellule e lega 4 ioni
Ca). Le proteine chinasi C vengono attivate dal Ca++,
dai digliceridi e dai prodotti di degradazione di alcuni fosfolipidi.
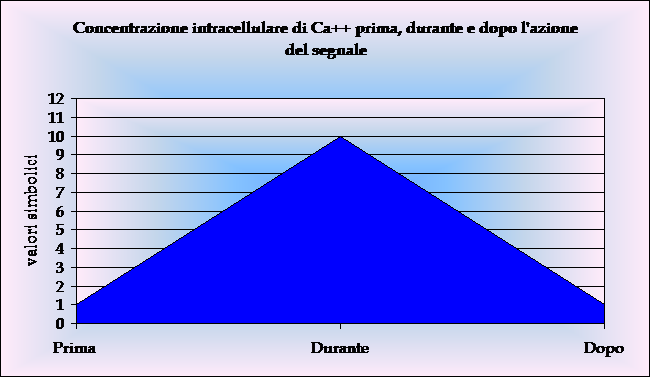
Grafico 2
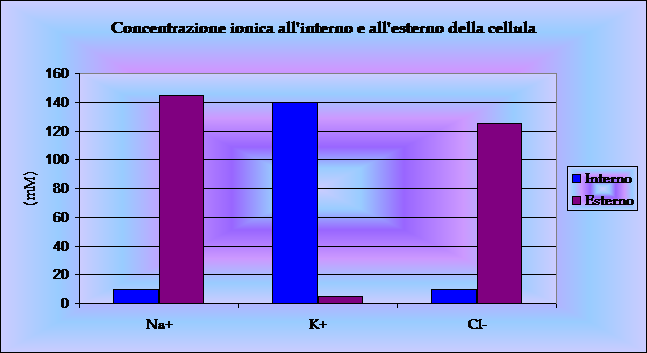
La gran parte degli ormoni e dei neurotrasmettitori, oltre che di altre
sostanze regolatrici, modificano i processi cellulari attraverso vie di
trasduzione del segnale nelle quali svolgono funzioni cruciali le
proteine leganti il GTP, denominate proteine G.
Dette proteine possono essere nello stato attivo o non attivo. Nel suo
stato attivo, le proteine G, hanno elevata affinità per il
GTP che
idrolizza in GDP, e fa ritornale la proteina allo stato inattivo.
Ricapitolando, il meccanismo di trasduzione del segnale mediato da una
proteina G e da una proteina chinasi, si compone di 6 fasi:
- una molecola regolatrice si lega al suo recettore plasmatico;
- il recettore occupato da detta proteina (il ligando), interagisce (attivandola) con una proteina G, la quale si lega al GTP;
- la proteina G attivata interagisce con uno dei secondi messaggeri inattivandolo;
- conseguentemente si verifica un aumento o un decremento cellulare di un secondo messaggero;
- l’aumento del livello di un secondo messaggero modifica l’attività di una o più proteine chinasi (dipendente dal secondo messaggero);
- si determina una variazione nel livello di fosforilazione di un
enzima o di un canale ionico che determina l’effetto finale.
L’adenilato ciclasi,
l’enzima che produce AMPc, è stato il secondo
messaggero scoperto per
primo. Tale enzima è sottoposto a controllo positivo e
negativo da
parte della proteina G. Se un ligando stimolante (es.: adrenalina) si
associa ad un recettore, causa l’attivazione di proteine G da
parte del
recettore stesso. La subunità alfas della proteina G
interagisce con
l’adenilato ciclasi attivandolo. Se invece una sostanza
regolatrice ed
inibente dell’adenilato ciclasi (es.: adenosina) si lega al
suo
recettore, attiva proteine G aventi una subunità alfa di
tipo
differente (alfai), in grado di inibire l’attività
dell’adenilato
ciclasi.
Fin qui abbiamo esaminato i casi in cui vari canali ionici sono modulati da agonisti extracellulari. Ma esistono anche canali ionici regolati da secondi messaggeri nei quali le proteine G rappresentano elementi fondamentali nel secondo stadio della cascata di trasduzione del segnale, altri ancora sono modulati in maniera diretta da proteine G, senza l’ausilio di secondi messaggeri. E’, ad esempio, il caso del legame acetilcolina-recettoreM2 (presente nel cuore). Tale legame attiva una proteina Gi la cui subunità alfa interagisce direttamente con una classe di canali del K+ aumentandone la probabilità di apertura.