
Questo termine deriva dal greco pteron ala e phiton
pianta, sono anche dette crittogame vascolari.
Sono piante aplodiplonti eteromorfe, con predominanza netta dello sporofito sul gametofito, che può assumere dimensioni molto ridotte. Sono caratterizzate da un’alternanza di generazioni, un’aploide, rappresentata dal gametofito e una diploide rappresentata dallo sporofito.
Il ciclo ontogenetico, ha inizio dalla germinazione
della spora che origina il gametofito o protallo, rappresentato da una
piccola lamina verde, che porta gli organi sessuali maschili o anteridi e
gli organi sessuali femminili o archegoni. Questi origineranno i gameti
maschili o anterozoi e i gameti femminili o ovocellule. Gli
anterozoi, sono in grado, grazie alla presenza di ciglia, di muoversi
nell’ambiente acquatico e penetrare nell’archegonio e fecondare in tal modo
l’ovocellula.
Questa, fecondata, originerà lo sporofito che si
differenzierà in fusto, foglie, radici.
Nelle pteridophyta, la presenza dell’acqua è
ancora essenziale perché possa avvenire la fecondazione, per questo gli
anterozoi sono provvisti di ciglia, che permettono il movimento nell’ambiente
acquatico.
Le foglie, di varia forma secondo la specie, portano
nella pagina inferiore gli organi di riproduzione asessuale, rappresentati dagli
sporangi, racchiudenti le spore, oppure da sori che possono o no essere protetti
da una membrana, detta indusio.
Il gametofito può essere tuberiforme, filiforme,
cordato, nastriforme, digitiforme. Il tuberiforme è molto arcaico. In genere ha
rizoidi, ma può anche esserne privo; in questo caso possiede internamente dei
funghi endofitici che provvedono ad assorbire l’acqua dal terreno. Solitamente
è autonomo, verde perché clorofillico. Importante è, che è il gametofito a
portare anteridi e archegoni.
L’anteridio è più o meno sferico e nel suo
interno produce gli spermatozoi o anterozoi; l’archegonio ha una parte
slargata o ventre in cui è situata l’ovocellula o oosfera e a cui fa seguito
una parte più allungata, detta collo, che sporge dal gametofito.
Gli anterozoi possono essere biflagellati o con un
ciuffo notevole di ciglia, come nell’Equisetum. L’archegonio, fecondato in
mezzo liquido, produce lo zigote, dapprima come cellula solitaria, che poi si
segmenta in due, in senso trasversale, parallelamente alla lamina del gametofito
(da ricordare che il collo dell’archegonio è rivolto verso il basso).
Di queste due cellule, la più bassa è detta
ipobasale, l’altra epibasale. La cellula ipobasale darà il sospensore col
compito di spingere verso l’alto la cellula che darà poi il corpo del
gametofito, a funzione protettiva; è sempre questa che darà il piede e magari
anche la radice che è accessoria, perché non si prolunga lungo l’asse
dell’embrione.
La cellula epibasale dà origine all’apice
vegetativo, cioè ad un fusticino minuto e ad un numero vario di cotiledoni. Si
segmenta sino ad avere una serie di gruppi cellulari, ognuno formato da otto
cellule e questo sino alla formazione completa dell’embrione, che vive a spese
del gametofito, in certo senso parassitandolo e rendendosi indipendente solo
quando emette le radici.
Ora l’embrione è costituito da apice vegetativo,
foglioline, cotiledoni, radichetta. Da adulto le “foglie” sono più numerose
e diconsi più propriamente fronde, in quanto sono foglie portanti gli apparati
sessuali.
Queste fronde possono essere piccole (microfillata) o
grandi (macrofillata). Le Licopodiaceae sono microfillata, l’Adianthus è
macrofillata. Interessante da notare è che nelle felci, le fronde portano,
nella parte sottostante, le parti destinate a dare le spore e il relativo
complesso di sori e sporangi, per questo si dà il nome di sporofilli,
alle fronde delle felci.
Accanto a queste, troviamo anche fronde senza spore
dette trofofilli, che sono sterili e deputate a nutrire la pianta. Gli
sporangi sulle foglie sono sempre ben visibili, oltre che nelle felci, anche
alla base delle piccolissime foglioline del Licopodium, sottoforma di piccoli
puntini gialli.
Sporangio è il concettacolo delle spore; se più
sporangi sono raggruppati insieme, essi costituiscono un soro. Lo sporangio può
essere ortotropo, se il suo asse è parallelo a quello del supporto; se invece
è pendente, è anatropo; mentre il primo non ha bisogno di un grosso
supporto, il secondo sì, perché deve essere sostenuto. All’interno dello
sporangio troviamo una zona detta tappeto che serve per nutrire le spore.
A volte, pezzi di questo tappeto si staccano con la spora e costituiscono
attorno ad essa il perisporio o perina, responsabile diretta della
superficie scolpita di molte spore
Più sporangi possono essere attaccati ad un asse
unico, centrale e costituire quell’entità detta strobilo; attorno
all’asse centrale dello strobilo, le varie entità fertili si dispongono a
spirale.
Se gli sporangi sono inseriti al margine o sotto la
fronda, parliamo di sporofilli; più sporangi possono raggrupparsi in una massa
e dare i sori, parola che significa semplicemente mucchio. Le piante possono
essere isosporee o eterosporee ed avere cioè, spore uguali o diverse; abbiamo
anche un’eterosporia fisiologica come nell’Equisetum, dove le spore sono
tutte uguali fra loro, almeno apparentemente, solo che alcune poi, daranno il
gametofito maschile e altre il gametofito femminile.
Ciascuna spora è fatta da un involucro esterno o esina
e uno interno o endina, spesso pluristratificato e talvolta ci può
essere anche, interno, il residuo della perina. La cellula madre della spora,
dividendosi, può dare origine a diversi tipi di spore; se disposte attorno ad
una linea centrale, a 90° le une dalle altre, sono spore monolete (come
gli spicchi di una mela tagliata in quattro); oppure abbiamo quattro spore
convergenti verso un centro puntiforme, con tre linee di contatto le une con le
altre. Sono spore trilete, a 120° le une dalle altre (4 tetraedri con la
base curva accoppiati). È importante questa distinzione, perché spesso
caratterizza tutti gli esemplari di un certo ordine. Il tipo trilete è il più
antico e, infatti, lo troviamo nei Licopodi, mentre fra le felci, la Dryopteris
ha il tipo monolete. Sono poi le monolete a presentare il residuo di tappeto,
cioè il perisporio.
Per l’antichità, vale quanto detto parlando della legione; abbiamo cioè l’Aldanophyton come pianta più antica; le più antiche sono Lycopsida e Rhyniopsida, già sicuramente presenti nel Siluriano; nel Devoniano abbiamo poi già quattro linee filetiche ben distinte.
Le affinità sono evidenti con Angiosperme, Conifere
e muschi. Possiamo dire che dopo la prima dicotomia, un ramo va alle felci e
l’altro si divide ancora in due, dando da un lato le Rhyniopsida e
dall’altro le Sphenopsida. Qui dovremo inserire le Lycopsida e le Filicopsdida.
Ciò che si conclude è, che è un problema molto complesso, Alcuni tendono a
fare degli Equiseti un gruppo separato. Hanno certamente un antenato comune, ma
non la Rhynia, dato che quando queste compaiono le Lycopsida sono già molto
sviluppate, inoltre le Rhynie non hanno lasciato discendenti. C’è
l’antenato comune, forse in un essere tipo Rhynia, ma non certo la Rhynia che
noi consideriamo oggi come linea filetica in parte a se stante. Alcuni ancora,
riuniscono in un solo gruppo Lycopsida e Sphenopsida. Qui si considerano
separati, per cui abbiamo quattro linee filetiche distinte.
Sono molto antiche, enormi fossili di 20-25 m.
d’altezza, con tronchi grossi. Il fusto non è articolato e i nodi sono
portati su una spirale; anzi spesso abbiamo due linee a spirale fra i vari nodi,
lungo il fusto. Anche le altre entità, oltre alle foglie, hanno questa
disposizione elicoidale.
Le foglie sono molto piccole e strette, terminanti a
punta nelle viventi e in genere biforcate nei fossili. Le dividiamo in due
categorie:
- le più antiche, dagli apparati sporigeni, detti
trofofilli.
- le più moderne dette impropriamente sporofilli,
perché qui, gli sporangi non sono inseriti sulle foglioline, ma in
zona
ascellare.
Gli sporangi sono costituiti da un’entità
uniloculare, cioè hanno una sola cavità, più o meno ampia, a ferro di
cavallo. Di regola sono solitari e ciascun sporangio s’inserisce alla base
della propria fogliolina; l’asse dello sporangio corrisponde sempre a quello
della foglia.
Nelle piante più antiche, abbiamo uno strobilo con
un suo asse, su cui si attacca la fogliolina; all’ascella di questa si attacca
l’asse dello sporangio, con in cima lo sporangio.
Nel corso dell’evoluzione si mantiene la posizione
reciproca di asse, strobilo, fogliolina, solo che l’asse dello sporangio si
salda a quello della fogliolina, per una maggiore protezione. Successivamente lo
sporangio scivola verso il punto di inserzione della foglia, per accorciamento
del suo asse. È questo un esempio di slittamento filetico per una migliore e
maggiore difesa.
A maturità le spore si staccano, solo
eccezionalmente germinano dentro lo sporangio, fenomeno accompagnato da
modificazioni profonde.
Gli spermatozoi sono biflagellati o con più
flagelli, meglio detti allora, pluriciliati. Ciò importa ai fini della
filogenesi; le più antiche hanno due flagelli, le più moderne sono
pluriciliate.
Consideriamo le seguenti sottoclassi:
1) Protolepidodendridae: appartengono al
Cambiano, Carbonifero e sono scomparse dopo il Paleozoico, per cui sono esistite
da 440 a 200 milioni di anni fa. Appartengono a questa sottoclasse l’Aldanophyton
e la Baragawanathia.
Le foglie mancano di ligula, sono forcate, talora
spinescenti, ossia con indurimenti apicali. Gli sporangi, almeno quelli noti,
sono portati da assi saldati alla foglia. Hanno una protostele senza midollo.
Ricordiamo l’Aldanophyton, che, se datato bene, appartiene al Cambriano
superiore. (è stato trovato in Siberia). Ancora da ricordare è la
Baragawanathia, sicuramente del Siluriano, con foglie ben evidenti, più antica
della Rhynia, in cui però non abbiamo foglie, per cui la presenza di foglie è
carattere più antico dell’assenza. È una pianta fossile australiana, ben
conservata.
Ancora da ricordare è il Drepanophycus.
2) Lycopodiidae: dal tardo Paleozoico
arrivano sino a noi con un notevole numero di specie, che i “conservatori”
classificano in un solo genere e i “divisori” in più gruppi, almeno
quattro, in relazione al fatto che il Licopodio europeo ha gli strobili ben
evidenti, mentre nell’etiopico no.
Vivono nella tundra, sopra i 2000 m., nelle foreste
tropicali, pendenti dagli alberi.
Sono senza ligula, isosporee, con spermatozoi
biflagellati; plectostele con protoxilema esarco.
Il caule dello sporofito, non presenta nodi e
internodi; i microfilli possono essere sparsi o disposti in modo elicoidale,
raramente a verticillo. Gli sporofilli, sono disposti all’estremità del
caule, in verticilli. Gli sporangi, solitari, sono alla base della parte
superiore dello sporofillo o all’ascella.
3) Selaginellidae: comprendono piante di varie
dimensioni. Risalgono al Carbonifero – Cretaceo e arrivano sino a noi con 600
specie tropicali e 4-5 specie nostrane. Sono eterosporee, con spore maschili
molto piccole e femminili grosse. Hanno la ligula, piccola appendice
all’inserzione della foglia, con funzione assorbente. Gli spermatozoi sono
biflagellati.
Hanno protostele o aciclostele, esclusiva di questo
gruppo e totalmente asimmetrica. Le meristele sono sempre 2-3, raramente di più.
Sotto le fronde presentano dei rizofori, appendici dicotomizzate
con struttura intermedia fra la radice e il fusto; ancorano la pianta al
terreno e hanno funzione assorbente.
Sono considerabili come gli ultimi rappresentanti di
quelle entità dicotomizzate, le stigmarie, che caratterizzeranno le fossili
Lepidodendridae, ritrovate in una zona dell’Inghilterra, ancora in piedi, come
una foresta pietrificata.
Ricordiamo la Selaginella, che nelle specie apus e
rupestris, ha la macrospora che invece di staccarsi e germinare nel terreno,
germina attaccata allo sporofito e anche la fecondazione avviene sullo sporofito;
l’embrione stesso, per un po’ di tempo, si nutre a spese dello sporofito e
poi si stacca. Questi fenomeni, si possono considerare come il preludio della
condizione definitiva delle Angiosperme, solo che qui il gametofito è ben
protetto, mentre per la Selaginella è all’aperto.
4) Lepidodendridae: risalgono al Devoniano e
sono tutte fossili, eccetto il solo genere Isoetes, presente anche in Italia, e
il genere Stilides, raccolto alcune volte sulle Ande e quivi presente in colonie
limitatissime. Nel Carbonifero, costituiscono quelle imponenti foreste, che oggi
ritroviamo pietrificate; erano piante alte 20 m. a fusto colonnare, terminante
con una serie di dicotomie, ciascuna con all’apice gli strobili. Molti tronchi
fossili, si trovano anche nel Sahara.
Sono eterosporee (spore maschile e femminili hanno
dimensioni diverse) ed hanno uno strobilo con macro e micro sporangi. La ligula
è presente. La stele è una protostele piena, con una massa interna di xilema
ed un’esterna di floema. Gli spermatozoi sono pluriciliati, almeno per le
piante viventi, per i fossili nulla sappiamo.
Hanno caratteristicamente le stigmarie, che
sono una specie di radici biforcate all’apice; sono molto sviluppate in
larghezza, che ci porta a pensare che vivessero in acquitrini. Le foglie quando
cadono, lasciano una cicatrice caratteristica a forma di rombo. All’apice del
rombo, troviamo traccia della ligula e al di sotto la traccia del vaso
cribrolegnoso, poi troviamo quattro entità costituenti il parichnos, cioè
le tracce di un particolare tessuto con ampi spazi intercellulari, che si
ritiene essere fatto da cordoni aeriferi, per cui sarebbe confermata l’ipotesi
di una vita in acquitrini per queste piante. Questa entità rombica è anche
detta cuscinetto fogliare; le tracce del parichnos possono avere posizioni
varie, restando fissa solo quella reciproca delle quattro entità, a formare una
sorta di X. L’insieme dei cuscinetti, costituisce quella sorta di reticolato
che è a maglie d’ampiezza variabile e che ci dà un’idea di come fossero
inserite le foglie nel loro complesso.
Alle Lepidodendrales appartiene la Sigillaria,
provvista di stigmarie, di un tronco dicotomizzato, da cui pendono gli strobili
di forma ovoidale. Al di sopra di essi, si trova il complesso delle foglie.
Le Lepidocarpales possedevano una macrospora
chiusa in un macrosporangio a sua volta chiuso da tegumenti. Le entità sono
strettamente unite; è un qualcosa che prelude ad una difesa dell’ovulo; la
macrospora germinava restando all’interno, ma poiché non si è dimostrato che
la fecondazione fosse effettivamente interna, non si può parlare di seme, anche
se il gametofito resta attaccato.
Nelle Isoetales, il tronco della Sigillaria si
è raccorciato fino a dare la Pleuromeia; poi si è ancora raccorciato a dare la
Nathorstiana (Cretaceo inferiore) e poi ancora fino allo Isoetes attuale, simile
ad una comune erbaccia, dove il fusto è solo la parte nerastra terminale.
Contemporaneamente si è ridotta la stigmaria a zaffo nella Pleuromeia ed è
ancora più piccola nella Isoetes velata.
L’Isoetes, che risale al Cretaceo ed è giunto sino
a noi, ha un fusto molto ridotto, da cui dipartono diverse foglie che sono anche
sporofilli, provviste alla base di una piccola incavatura detta fovea. I
macro e i microsporofilli sono inseriti sopra la fovea. Possiede la ligula.
Sezionando troviamo traccia della ligula, del fascio cribrolegnoso, di sei
canali, di cui quattro auriferi e due mucipari, ultimo residuo del parichnos.
È l’ultimo esemplare vivente di un gruppo molto
importante del passato.
Le caratteristiche principali del gruppo sono: un
caule bulboso, sporofilli e trofofilli simili, con ligula presente e con cavità
ove ha sede lo sporangio che è ricoperto da una membrana, detta indusio.
Il protallo molto piccolo, è localizzato all’interno della spora da cui è
generato, è dioico in quanto le macrospore danno protalli femminili e le
microspore protalli maschili. Le lunghe radici hanno una divisione dicotomica.
Gli spermatozoi sono pluricigliati.
Le Miadesmiales vivevano nel Carbonifero ed
erano piccole, più o meno erbacee.
Sono piante molto antiche, con molti fossili; sono
giunte sino a noi con un solo genere e poche specie, presenti un po’ ovunque.
Tipico rappresentante, è l’Equisetum, che può essere considerato come un
fossile vivente, essendo tale e quale allora e risalente a più di due milioni
d’anni fa.
Hanno fusto articolato, diviso in nodi e
internodi, presenti anche nelle ramificazioni laterali. Dal nodo partono rami e
foglie che sono a verticillo, per cui siamo alla presenza di un sistema
multilacunare. I rami sono posti sotto le foglie ma su ortostiche diverse; le
ortostiche diverse sono in relazione al fatto che, altrimenti la pianta non
avrebbe più alcuna resistenza in quel punto e si affloscerebbe.
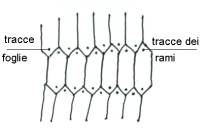
Le foglie sono piccole, per cui sono piante
microfillata, possono essere più o meno lunghe, ma sempre strette e al più
flagellate (foglie di Hyenia) e profondamente incise. Formano sempre uno o tre
verticilli oppure sei, nove, dodici, ma sempre multipli di tre nelle più
antiche.
Gli sporangi sono tutti anatropi, cioè il supporto
si ripiega ad uncino ed il peduncolo dello sporangio viene così ad essere
parallelo al sostegno. Interessante è la presenza di un lungo peduncolo che
permette il rovesciamento dello sporangio, ciò ha poi permesso una maggiore
difesa, fino ad aversi una sorta di scudo portante sotto gli sporangi.
Nella maggior parte dei casi c’è una sorta di
strobilo, cioè un asse principale con attorno gli sporangi; in altre piante
l’asse ha finito per saldarsi ad una foglia che protegge ora gli sporangi come
nelle Sphaenophyllales.
Potremmo chiamare questa brattea, sporofillo, ma è
sui generis, perché si è saldata all’asse dello strobilo. Sporangio,
peduncolo portante, formano un insieme inscindibile; più sporangi insieme per
dicotomizzazione del peduncolo, formano lo sporangioforo.
Le spore si staccano subito, per cui il gametofito si
forma sempre nel terreno; esse non germinano mai sulla pianta. Gli spermatozoi
sono provvisti di moltissime ciglia, da un solo lato.
Questo è un gruppo molto importante, risalente al
Paleozoico; esistono Sphenopsida colonnari, cioè a grosso fusto, incapaci però
di dare nuovi polloni; esistevano anche piante lianose o almeno molto deboli che
si aggrappavano e attorcigliavano attorno ai fusti. Questo si deduce dai
ritrovamenti fossili dei loro fusti che sono molto piccoli, sempre con un certo
grado di torsione.
Come probabili antenati si citano il Climaciophyton e
lo Spondilophyton, che però sono piante molto piccole e poco note, per cui i
dubbi sono molti e validi.
Si dividono in:
Hyenidae: risalgono al Devoniano medio. Caratteristica
fondamentale è che manca l’articolazione o è irregolare; anche le foglie, in
numero di tre, sono irregolarmente verticillate (Hyenia), mentre nel
Calamophyton, il fusto è articolato. In generale il fusto era bi-, tri-forcato
e anche le foglie erano più o meno profondamente divise. Siamo però di fronte
ad un’anisogamia, per cui le dicotomie non sono più uguali da ambedue le
parti.

Interessante è la situazione per gli sporangiofori.
Nel Calamophyton abbiamo un supporto che si divide apicalmente in due parti,
ciascuna di esse porta il suo sporangio; l’Hyenia presenta successive
divisione del peduncolo, per cui gli sporangi accoppiati sono almeno sei. La
moltiplicazione degli sporangi determina il moltiplicarsi dei rami secondari del
peduncolo.
Gli sporangiofori sono, nel complesso, organizzati a
costituire uno strobilo, cioè un asse centrale, cui sono inseriti, a spirale o
a verticillo, le parti produttive degli sporangi.

1 – Calamophyton
2 – Hyenia 3-
Archaeocalamites 4- Equisetum
5 – Lingularia
6 – Calamostachys 7 –
Paleostachys
Pseudoborniidae: sono rappresentative del Devoniano
superiore. Gruppo poco conosciuto, avevano foglie ripetutamente forcute, per
successive anisodicotomie. Gli sporangi sono organizzati in sporangiofori e fra
uno e l’altro di questi, compaiono delle brattee fogliari.
Sphenophyllidae: sono presenti dal Devoniano al Mesozoico.
Sono piante probabilmente volubili, lianose; le foglie in numero di tre o
multipli di tre, fino a dodici al massimo, flabellate, piccole, a forma di
ventaglio, a volte divise in due. Gli sporangi si sono saldati alla brattea
protettiva, ma l’asse dello sporangioforo è distinto da quello della
fogliolina; sono entità saldate fra loro, non fuse. Erano iso o eterosporee,
per cui gli strobili portavano micro e macrospore.
Anatomicamente presentavano, come struttura primaria,
una stele triarca, fatta da una massa centrale di xilema, con protoxilema
esarco. L’inserzione delle foglie avviene in corrispondenza dei “vertici del
triangolo”, sono quindi, al minimo tre e per successive divisioni possono
arrivare fino a dodici. Il fusto, nel corso del tempo, assume una struttura
secondaria, proliferando il deuteroxilema in corrispondenza dei tre vertici.
Lentamente si formano due anelli di floema e xilema. Questa stele è esclusiva
del gruppo.
Equisettidae: è il gruppo più importante, che dal Devoniano
superiore giunge fino a noi con l’Equisetum o coda di cavallo. I primi
rappresentanti sono diversi dal nostro che compare solo nel Carbonifero.
Il caule dello sporofito è articolato, con evidenti
nodi e internodi; sui nodi sono inseriti i rami verticillati, anche le foglie
sono disposte in verticilli alterni fra loro. Le cellule sono silicizzate.
Gli sporangiofori sono inseriti su di un asse a dare
una sorta di strobilo; possono mancare di parti intermedie fogliacee, per questo
motivo le dividiamo in due gruppi sistematici. Sono iso o eterosporee; possono
presentare eterosporia fisiologica, per cui spore uguali danno gametofiti
diversi.
Calamitales: presenti dal Devoniano al Triassico e
poi si estinguono. Le parti fogliacee sono intercalate agli sporangiofori per
cui lo strobilo è fogliaceo; abbiamo tre tipi di brattee fogliari. Nella
Cingularia, le foglie stanno sopra gli sporangiofori, nella Calamostachys sono
presenti anche due foglie, nella Paleostachys abbiamo una foglia sola. In realtà
abbiamo avuto uno slittamento filetico; la forma più primitiva era quella della
Cingularia in cui il ramo era considerabile quale sporangio, data
l’equivalenza delle due posizioni. Per gli altri due tipi, si ammette, o uno
spostarsi della brattea, o dello sporangioforo. Non si sa con precisione, ma si
conviene sia la brattea; essa si sposta verso l’alto e viene ad essere
intermedia o sottostante gli sporangi, realizzando così una difesa maggiore di
prima, perché protegge anche l’ultimo sporangioforo.
Equisetales: mancano di brattee intermedie difensive;
questo è in relazione al moltiplicarsi del numero di sporangi per sporangioforo
e dei relativi peduncoli di II ordine, anzi, tutti gli assi secondari, si sono
saldati insieme in una sorta di scudo od ombrello, che porta ora al di sotto
tutti gli sporangi. Gli ombrellini, tutti inseriti sull’asse centrale dello
strobilo, da giovani sono tutti a stretto contatto, si ha così una difesa degli
sporangi. Allungandosi l’asse dello strobilo contemporaneamente alla
maturazione delle spore, gli ombrellini si distanziano e si ha la dispersione
delle spore.
L’Equisetum è il solo rappresentante vivente; è
un genere con circa 20 specie, alcuni non molto grandi, altri alti fino a 3-4
metri, come quelli tropicali. Compare nel Carbonifero inferiore come Equisetites
e poi prospera, con esemplari del Mesozoico in tutto uguale ai nostri, fino a
noi.
Hanno fusto con lacuna centrale, per alleggerimento,
e distinguiamo tre tipi di stele, in relazione al comportamento dell’endodermide.
Nel I tipo troviamo lacune periferiche in corrispondenza alle coste del fusto,
dette lacune carinali, accompagnate da lacune vallecolari più
grosse ed in corrispondenza degli avvallamenti. I fasci vascolari sono
equidistanti dal centro, separati da raggi midollari ed è presente un’unica
endodermide. Nel II tipo compare una seconda endodermide, più interna; è
questo un raddoppiamento di entità, che ritroveremo anche nelle piante
superiori, in relazione alla presenza di cavità (fasci bicollaterali nelle
zucche, piante con cavità centrale). Nel III tipo, abbiamo un’endodermide
unica, ma divisa e ogni fascio e lacuna carinale, si circonda di un proprio
endoderma. Il I tipo è definibile quale eustele; il III tipo come dictiostele;
il II tipo, è considerato come termine di passaggio, dato che le due
endodermidi sono festonate in senso opposto.
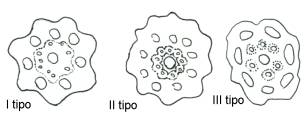
Hanno un corredo cromosomico di 108 cromosomi, numero
alto, derivante da poliploidia che si conserva regolarmente in tutti gli
individui.
Le spore sono di tipo particolare; oltre ad avere
l’esosporio e l’endosporio, comuni a tutte le spore, ne possiedono un terzo,
l’episporio che quando la spora è matura si divide in quattro “nastri”
detti apteri o amplessori. Queste appendici sono igroscopiche, e
in ambiente secco sono arrotolate alla spora, in ambiente umido si aprono
distendendosi e aiutano la spora a fissarsi nel terreno per poter germinare.
Dalle spore nascono gametofiti diversi, si ha quindi eterosporia fisiologica.
Gli anteridi sono portati alla sommità del gametofito maschile, mentre l’archegonio
è completamente infossato. Gli spermatozoi hanno un ciuffo di ciglia apicali.
Lo sporangioforo è fatto dalle parti apicali saldate in un pileo o scudo; i
vari scudi sono da giovani ravvicinati alla sommità del gametofito femminile,
poi per un allungamento dell’asse, si allontanano e le spore possono
disseminarsi.
Per i rami valgono le regole generali: sono inseriti
sotto il punto d’inserzione delle foglie.
Si conoscono 25 specie presenti in tutti i
continenti, ad eccezione dell’Australia.
Ricordiamo l’Equisetum arvense o coda di cavallo,
presente nei luoghi umidi, l’Equisetum fluvialis, l’Equisetum hyemale, l’Equisetum
palustre.
L’Equisetum arvense è il più tipico, ha un rizoma
sotterraneo molto sviluppato e, in primavera sviluppa piante fertili, di piccole
dimensioni, di colore bruno, non ramificate, terminanti con una spiga che
racchiude gli sporangi. In estate, sviluppa steli sterili, alti sino ad un metro
e con rami uniti a verticillo che fuoriescono dai nodi del fusto.


Sono tutte fossili. Sono state aggregate ai gruppi più
disparati, di esse si conosceva solo la parte sterile e così furono messe
insieme alle Cycas; dopo la scoperta della parte fertile, sono state inserite
fra le Pteridophyte, cosa migliore è isolarle.
Per l’inserzione delle foglie, ricordano le
Sphenopsida e pur non avendo il fusto articolato, hanno le foglie inserite su
ortostiche diverse.
Gli sporangi sono molto numerosi, inseriti
all’apice del fusto, sulla pagina superiore della foglia trasformata. Sono
eterosporee. I macrosporangi sono inseriti su squame che s’incastrano una con
l’altra, formando così un ambiente protettivo. Gli strobili sono di tipo
particolare, perché mentre nelle Sphenopsida c’è una disposizione a
verticillo e nelle Lycopsida una disposizione a spirale, qui invece, sono su due
o quattro ortostiche. Questa disposizione è stata vista con difficoltà e solo
nel 1915 si vide questa rispondenza nella disposizione delle parti fertili e
delle sterili.
Si suddividono in due sottoclassi: Noeggerathiidae
e Tingiidae.
Le Noeggerathiidae risalgono al Carbonifero e vanno
fino al Permiano; le Tingiidae vanno dal Permiano al Triassico. È difficile
ammettere una discendenza diretta delle une dalle altre.
Le Noeggerathiidae hanno due serie di foglie,
uguali fra loro e disposte in due file diverse; la stessa disposizione è per
gli strobili, ogni pinna è una foglia, l’insieme delle foglie è sormontato
dagli apparati microsporangiali, caratterizzati dal gioco d’incastro delle
diverse unità.
Le Tingiidae, che sono rinvenibili solo in
Asia, hanno le pinne disposte su quattro fila, di diversa dimensione. Le bratte
stesse, sono entità strette, portanti quattro sporangi per ciascuna brattea;
qui si potrebbe pensare ad una fusione o saldatura di più sporangi fra loro.
Ogni Tingiida, possedeva solo due strobili, appaiati su rami biforcati.
L’avere le foglie disposte su quattro fila, cioè su due piani fra loro
perpendicolari, fa pensare ad una sorta di verticillo, che le riavvicinerebbe
alle Sphenopsida; gli sporangi su brattee, ricordano le Sphenofillales, ma con
la grandissima differenza, che là gli sporangi sono anatropi, qui ortotropi; là
abbiamo saldatura degli sporangiofori alla costa centrale della lamina fogliare,
per cui non c’è uno sporofillo, mentre qui, si ha un vero sporofillo.
D’altra parte, non possiamo per questo mettere le Noeggerathopsida fra le Felci, perché fra le Felci, gli sporangi sono sulla parte dorsale delle foglie, qui sulla ventrale; non si può ammettere una rotazione degli apparati, perché questa avviene lentamente e raramente.
Pochi sono i rappresentanti viventi e solo due le famiglie: le Psilotaceae e le Tmesipteraceae. È da distinguere nettamente dalle Psilophytopsida, nome un tempo usato per indicare quelle che oggi sono le Rhyniopsida e poi abbandonato per evitare confusioni, dato il suono similare.
Non conosciamo fossili. Il fusto non è articolato e i fusticini sono diritti con una dicotomia o con una serie d’isodicotomie, sfasate fra loro di 90°; all’interno del caule è presente un piccolo vaso conduttore assiale. Le foglie sono piccolissime in Psilotum, più grandi in Tmesipteris, in relazione al fatto che nel primo non sono vascolarizzate e nel secondo sì. Lo Psilotum, manca pure di stomi. Sporofilli e trofofilli sono diversi, i primi fertili, sono biforcati.
Gli sporangi sono riuniti a dare dei sinangi, termine con cui intendiamo la fusione di sporangi, che in Tmesipteris sono due entità e in Psilotum tre.
Ciascuno sporangio ha un proprio tessuto vascolare e le varie entità sono unite, ma al centro hanno una colonnina nutriente. Non danno strobili, ma sono solo portati dal fusto. Fino ad ora era stato sempre presente un asse centrale portante le diverse unità, questo è il tipo più primitivo da cui deriveranno i tipi con disseminazione lungo il fusto e più tardi, quelli lungo la foglia.
Lo Psilotum è una pianta molto caratteristica con rizoma e gametofito uguali, per cui li riconosciamo solo se il primo ha apparato radicale e il secondo anteridi e archegoni. Questo gametofito ha accrescimento indefinito, manca di rizoidi e vive in simbiosi con funghi endofitici (micorriza). In certi fossili di zone vulcaniche, il gametofito è vascolarizzato. Il sinangio è fatto da tre entità. Esso è accompagnato, alla base, da due piccole squamette che interpretiamo come foglie, ma non stomi né vascolarizzazione. Nella Rhyniopsida, il sinangio è fatto da due entità unite, accompagnate da squame molto lunghe e vascolarizzate. La differenza fra le due famiglie di Psilotaceae e Tmesipteraceae è il numero degli sporangi per sinangio, nelle squame e nella vascolarizzazione.
Le Psilotopsida sono assai vicino alle Rhyniopsida e anzi alcuni le considerano come gli ultimi discendenti. Me nelle Rhyniopsida non abbiamo appendici fogliari né sinangi apicali; inoltre sono già estinte all’inizio del Carbonifero e le conosciamo solo come fossili.
Psilotum e Tmesipteris non hanno fossili ed è difficile ammettere una discendenza non conoscendo la catena d’esseri intermedi.
Le Rhyniopsida danno fossili nel Devoniano, con struttura
conservata anche del citoplasma, quindi ci si chiede come non abbiano dato più
fossili in epoche successive, se così bene fossilizzavano.
Dette Psilotophytopsida e poi diversamente battezzate
per evitare confusione, sono tutte fossili. È un gruppo comparso nel Siluriano
e scomparso nel Devoniano.
La loro scoperta risale ai primi del secolo, quando
furono trovate, ben conservate, in Gran Bretagna. Consideriamo la Rhynia più
primitiva e l’Asteroxilon più evoluto.
Il fusto non è articolato, raggiunge i 45 cm. nella
Rhynia, un metro in Asteroxilon. Il fusto ha una parte sotterranea, circa ad
angolo retto con la parte superiore, senza radici, tuberiforme, per cui si pensa
esistessero funghi endofitici simbionti. Incerta è l’interpretazione del
rizoma.
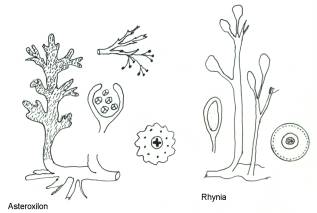
Il fusto è portato dal rizoma e le ramificazioni
erano completamente afille nella Rhynia, o con numerose appendici in Asteroxilon,
non ci sono foglie ma emergenze spinose; sono presenti gli stomi e sembrerebbe
fossero verdi.
Gli apparati sporigeni sono apicali e in Asteroxilon
su rami ben distinti. Le spore sono tetradi, cioè quattro entità unite
insieme, ciò fa pensare che la meiosi non portasse alla separazione delle
quattro entità, era cioè una meiosi imperfetta e di queste entità una sola
germinava e le altre servivano probabilmente per nutrimento. Erano spore trilete,
con tre linee di contatto fra le entità, ulteriore conferma che il tipo trilete,
già visto nel Lycopodium, era più primitivo dell’altro, monolite, a quattro
spicchi di mela. Notevole in questi fossili l’evidenza del floema. La stele è
una protostele stellata, con coste e valli; tutta la massa xilematica è intera
senza midollazione, di tipo primitivo, in corrispondenza con i caratteri arcaici
del gruppo. Hanno endodermide ed epidermide e tessuti parenchimatici tipo canali
aeriferi, in relazione a una loro probabile vita in paludi. Prima della scoperta
dello Psilotum, si parlava di un rizoma con parte basale ed aerea, quando si
vide che lo Psilotum vulcanico della Nuova Zelanda aveva il gametofito
vascolarizzato, il rizoma fu detto gametofito e la parte aerea erano i rami
sporofitici che da esso partivano. Venivano perciò ad avere un termine di
passaggio fra muschi e felci, perché la parte sporofitica viveva a carico del
gametofito ma era vascolarizzata. Era così un gruppo intermedio. Il fatto è
stato molto discusso, ma per accettarlo senza riserva, sarebbe necessario
trovare un fossile con residui di archegonio e anteridio sul gametofito-rizoma,
ma non è mai stato trovato alcunché del genere. Per ora, quindi, ci conviene
ammettere che il tutto è sporofito, anche perché di sicuro si sa poco.
Talvolta troviamo anteridi e archegoni sul rizoma, ma senza rami e quelle che
erano considerate come due specie con gametofito e sporofito distinti, sono in
realtà una sola specie, con gametofito e sporofito insieme sulla stessa pianta.
Anteridi e archegoni sono di tipo pteridofitico, per
cui non possono essere considerati anelli di congiunzione fra muschi e felci.
Rhyniidae: la Rhynia, è il tipico esemplare per la teoria del
teloma, secondo cui il fusto è la parte fondamentale e le foglie sarebbero nate
per una coalescenza dei rami del teloma.
Ha fusto afillo, sporangi terminali, con spore
interne, sparse.
La Rhynia avrebbe perso le foglie nel corso
dell’evoluzione, non come condizione primigenia, infatti, si sono trovati
fossili più antichi e completamente fogliati.
Nell’Horneophyton c’è invece nello sporangio,
una columella interna, considerata la prova dell’origine di questa classe dai
muschi.
Ricordiamo ancora la Yarravia del Siluriano,
singolarmente provvista di sinangi, fatto che la ravvicina alle Psilotopsida.
Psilophytiidae: afille, con fusto sotterraneo e sporgenti
dal terreno solo con rametti spinosi; gli sporangi erano portati nella parte
terminale senza spine.
Asteroxylidae: il fusto di Asteroxylon è completamente fogliato,
ma all’apice, i rami sono senza foglie e portano gli sporangi. Questi hanno
all’apice un piccolo forellino, verso cui confluiscono degli ispessimenti
cellulari, in una sorta di primo abbozzo di un apparato di deiscenza che avrà
maggiore sviluppo nelle felci. Le spore sembrano trilete, infatti ci sono
quattro entità fossili che non si sa come fossero organizzati fra loro; i
rametti sporigeni non sono mai stati trovati attaccati all’Asteroxylon.
È un gruppo assai vasto, presenti soprattutto nei
paesi tropicali; si ritengono esistenti circa 15.000 specie note, di cui solo 60
europee. I nostri rappresentanti sono piccoli, erbacei, mentre nei paesi
d’origine, le felci sono alte 6-7 m., la Cyathea raggiunge i 20 m. d’altezza
e 50 cm. di diametro.
Il fusto dello sporofito, ipogeo, non è mai
articolato, cioè non ha nodi né internodi; le fronde, trofofilli (senza
sporangi e a funzione trofica) e sporofilli (portanti gli sporangi), sono uguali
fra loro. Anatomicamente ricordano più un ramo che una foglia, sono ampie,
perciò sono piante macrofillata. Il gametofito o protallo, generalmente
monoico, può assumere diverse forme, con rizoidi e può o no essere
clorofillico.
Gli sporangi sono a grappolo (Osmunda), lineari (Asplenium),
puntiformi (Polipodium) ecc. Sono fatti in modo diverso da pianta a pianta; in
genere abbiamo più sporangi riuniti insieme a costituire i sori che, possono
essere portati sulla pagina inferiore o dorsale della foglia, al margine, al
centro, lungo le nervature; qualora costituiscano grappoli, sono sempre disposti
lungo una foglia che ha avuto una contrazione delle parti laminari. La forma e
la disposizione degli sporangi, ha carattere diagnostico. Le spore si staccano
prima di germinare; talvolta si stacca tutto lo sporangio, talvolta si ha
germinazione dentro il macrosporangio. Gli spermatozoi hanno numerose ciglia, a
volte elicate. Schematicamente, possiamo rappresentare il ciclo ontogenetico con
la spora che germinando origina il gametofito o protallo. Esso porta anteridi e
archegoni. Gli spermatozoi, prodotti dagli anteridi, penetrano, in presenza
d’acqua, negli archegoni e fecondano l’oosfera. Lo zigote formatosi, come
prodotto della fecondazione, dà origine all’embrione che si svilupperà in
sporofito sfruttando il protallo. Sulle foglie dello sporofito, si formeranno
gli sporangi entro i quali si sviluppano le spore. Il gametofito rappresenta la
fase aploide, lo sporofito, predominante, la fase diploide.

Le spore germinano dando ciascuna il gametofito
giovane, che poi diventa adulto, ed è rappresentato da una piccola laminetta di
2-3 mm., cuoriforme, plagiotropo, con rizoidi e parte femminile e maschile. In
genere la parte femminile sta nell’insenatura a cuore, la maschile più in
basso vicino ai rizoidi. L’archegonio è fatto da una parte basale o ventre,
inserita dentro il gametofito e proseguente col collo, allungato e sporgente.
Nel ventre c’è l’oosfera. L’anteridio, più o meno ovoidale, con parete
sottile, contiene i gameti maschili o spermatozoi (anterozoi), provvisti di
sfera basale, appendice elicoidale con ciglia terminali.
Quando nell’ambiente c’è acqua a sufficienza, si
rompe e gli spermatozoi fuoriescono; attratti dall’acido malico prodotto dal
collo dell’archegonio, entrano in esso e fecondano l’oosfera, formando ora
lo zigote. Esso si divide in due cellule, l’ipobasale e l’epibasale, da cui
deriverà l’embrione, che vive ora a spese del gametofito, cui è attaccato
col piede. Quando si è accresciuto abbastanza, emette una radice, la prima
fronda e diventa indipendente. Si sviluppa ora la giovane pianta che si
accrescerà fino allo stato adulto. Veniamo così ad avere una fronda fertile,
che si trova attaccata ad un rizoma sotterraneo, rappresentante il fusto. Questa
fronda, a maturità produrrà gli apparati sporigeni sulla pagina inferiore.
Ingranditi appaiono come piccole laminette reniformi. Ciascuna di esse è un
soro, cioè un complesso di sporangi. Il soro è ricoperto da una membranella
detta indusio, che può anche mancare, secondo i generi. Dentro troviamo gli
sporangi; nello sporangio, hanno origine le spore. In esso, infatti, abbiamo un
archesporio che si moltiplica, dando le cellule madri delle spore, ciascuna
delle quali, per meiosi, da quattro, o multipli di quattro, spore. Qui lo
sporangio è fatto da pareti molto tenui, unistratificate, che nella parte
terminale portano un anello di cellule ispessite. L’anello non è continuo,
perché interrotto dall’inserzione del peduncolo e dal punto di rottura
prestabilito detto stomio.
All’interno dello sporangio troviamo il tappeto
che nutre le spore e il cui residuo costituisce il perisporio o perina. Lo
sporangio è provvisto di un anello, fatto da una fila di cellule con parete
tangenziale ispessita ad U ed una parete più sottile alla sommità della U, con
all’interno una gran quantità di citoplasma. Con la maturazione delle spore,
le cellule dell’anello disseccano, e così tendono a congiungersi, facendo
pressione verso l’esterno; le spore fuoriescono e il ciclo ricomincia.
Quest’ultimo fenomeno è detto eversione dell’anello.
Complessa è l’anatomia delle felci; abbiamo,
infatti, protostele, protostele stellata, sifonostele, dictiostele, sifonostele
anfifloica, polistele, policiclodictiostele; abbiamo cioè, un’infinità di
tipi di stele. I più comuni sono sifonostele e dictiostele che si possono
considerare derivate da un tipo primitivo che per midollazione ha dato i due
tipi con cilindri concentrici di floema e xilema interi, dapprima e poi
spezzettati in tante meristele. Come meristemi né abbiamo una varietà molto
grande. Quando dal fusto a sifonostele escono le tracce fogliari che daranno la
fronda, in definitiva è un pezzetto di sifonostele che sporge fuori e migra
nello stipite (picciolo della fronda), formando la parete vascolare della fronda
stessa. Se sezionassimo uno stipite, troviamo endodermide, floema, xilema, cioè
una zona corticale con endodermide, periciclo e dentro floema e xilema. Anche
l’endodermide migra nella foglia e circonda la meristele, cioè la porzione di
stele che è migrata. Nella dictiostele è addirittura un’intera meristele che
migra nello stipite, anche qui con l’endodermide; da cui si deduce che la
fronda è qualcosa di più simile ad un ramo che ad una foglia. Svariatissima è
la forma delle pareti vascolari all’interno dello stipite; abbiamo tipi a C,
X, H, V, U, T, ciò ha valore diagnostico e ci permette di riconoscere un genere
dalla sezione. Nello stipite, possiamo avere due fasci con la loro endodermide.
Nello stipite di Pteridium aquilinum, una felce abbastanza comune, troviamo
anche 20 meristeli, derivanti da un’iniziale, che si è suddivisa;
nell’insieme le meristeli danno una sezione a omega.
La classificazione è difficile, perché è un gruppo
molto antico che ha enormemente prolificato fino a noi, possiamo asserire che
l’evoluzione sia ancora in atto.
Modernamente, lo studio dei cromosomi ha portato un
contributo insperato, perché mantenendosi costante il loro numero, si possono
determinare famiglie ben definite a 13n, 29n, 31n, e altre. Questi studi
cariologici hanno confermato l’alto numero di famiglie.
Possiamo classificarle in base alla posizione dei
sori, ma anche alla loro origine, da cui, la divisione in Eusporangiate,
se gli sporangi derivano da un gruppo di cellule e in Leptosporangiate,
se gli sporangi derivano da una cellula unica. Si è visto, però, che entità
eusporangiate possono essere leptosporangiate, per cui si va pian piano
abbandonando detta divisione. Si è parlato anche di Hydropterideae, come gruppo
a sé, ma lo stare in acqua non è carattere sistematico così differenziato. Le
dividiamo in diverse sottoclassi, rispecchianti l’antichità dell’origine.
Hanno fronde composte di un insieme di assi su piani diversi, così da costituire una serie di ramificazioni dicotomizzate e anche tricotomiche. Gli assi sono posti in piani diversi da quello del supporto, per cui i diversi piani s’intersecano. È questa una disposizione strana ma interessante ai fini della teoria della cladodificazione; questo, perché la precisa disposizione in più piani, può fare pensare che si sia avuta cladodificazione, che però, ha interessato solo alcuni piani, per successivo raccorciamento e contrazione dei rami stessi, che nel corso dell’evoluzione si sarebbero organizzati al massimo solo su tre piani.
Hanno appendici molto evidenti, dette aflebie,
mancanti di tessuto vascolare e che oggi ritroviamo in alcune piante viventi, ma
solo come appendici rudimentali. Gli sporangi erano terminali, ben distinti,
sferici, aprentisi per la fuoriuscita delle spore, con pareti pluristratificate.
Si estinsero molto presto, durando solo dal Devoniano medio al Permiano; gli
unici loro discendenti potrebbero essere le Ophioglossidae, ma è incerto.
Ricordiamo Arachnoxylon, Iridopteris, Botryopteris,
Stauropteris, alcune iso- e altre etero-sporee. Si è giunti alla conclusione
che questo è un gruppo molto complesso; forse i generi eterosporei non
sarebbero felci, ma una linea filetica a sé che potrebbe essere l’antenata
delle Pteridospermopsida.
Ai tropici sono rappresentate con 3-4 generi, da noi
esiste l’Ophyoglossum vulgare e l’Ophyoglossum lusitanicum, presente
nell’isola di Linosa. Non abbiamo fossili, ma per le loro caratteristiche,
sono sicuramente arcaiche.
Hanno prefogliazione diritta e non circinnata, come
normalmente, fatto eccezionale; le parti giovani sono chiuse da guaine che si
aprono quando spunta la foglia; la parte fertile è una spiga o un complesso di
spighe; la parte sterile è una lamina a mo’ di foglia.
Gli sporangi non hanno l’anello, ma un punto di
rottura netto, senza una struttura deiscente differenziata; la parete è
pluristratificata, carattere arcaico; sono eusporangiate.
Il gametofito è più o meno cilindrico, tuberiforme,
allungato e bitorzoluto, con accrescimento indefinito; talvolta è quiescente
così a lungo da dare gli apparati riproduttivi solo ogni dieci anni. Sono
piante micorriziche e anzi alcuni pensano che la quiescenza della pianta sia da
mettersi in relazione ad una quiescenza da parte del fungo endofitico; si
avrebbe lo sporofito solo quando sono “svegli” entrambi.
Sono piante viventi dal Carbonifero superiori ad
oggi. I generi di allora, erano diversi dagli odierni. Le fronde hanno
prefogliazione circinnata e le pinne misurano sino ad un metro e mezzo
d’altezza. Alla base della foglia, troviamo due grandi stipole che riparano la
fronda e si aprono come due grandi conchiglie protettive. Hanno canali lisigeni
con abbondante mucillagine. Sono tropicali e di difficile coltivazione in serra
perché vogliono un’umidità del 100%. Vivono in vicinanza dei ruscelli. Hanno
sporangi eusporangiati, senza indusio, senza anello, distinti o fusi in sinangi
a sfera o a rosetta.
Compaiono nel Permiano e giungono sino a noi. È
sicuramente una linea conservatrice, rappresentata da piante che vivono in
presenza d’acqua, non sono molto comuni e si suddividono in tre generi. Hanno
una rappresentazione cromosomica di 2n=22 per cui n=11, numero molto basso,
singolare omogeneità cromosomica, carattere d’accordo con altri fatti
convergenti verso la linea conservatrice. Fossile dell’ultimo paleozoico e
simile agli attuali è l’Osmundides
Gli sporangi sono di tipo intermedio fra
Eurosporangiate e Leptosporangiate. La prefogliazione è circinnata, le fronde
possono essere pelose, talvolta ghiandolari, con parte fertile e sterile insieme
o no. Certe fronde di Leptopteris sono a tre soli strati di cellule, qualcosa di
simile ai filloidi dei muschi. Gli sporangi hanno i primi cenni di un apparato
di deiscenza con cellule periferiche a pera, ispessite, deiscenti, ma senza
anello. Le spore sono autonome, in quanto hanno la clorofilla.
Il gametofito è perenne, appiattito, più o meno
simile a quello delle Filicidae.
È il gruppo più complesso è più rappresentativo.
Sono piante molto diverse fra loro, il che giustifica la divisione in tante
famiglie. Sono del tardo Paleozoico e giunte fino a noi, con dapprima un periodo
di sviluppo limitato e poi abbondante prolificazione nel terziario, dando più
di 12.000 specie.
La prefogliazione è circinnata, sono prive
d’appendici stipuliformi, ma spesso alla base delle fronde, troviamo peli o
appendici squamiformi, dette palee. Sono importanti per il significato nel
gruppo delle felci, in quanto importante carattere diagnostico per il genere e
per la specie. Talvolta invece delle palee, abbiamo peli di 3-4 cm. a dare un
rivestimento giallino. Sono leptosporangiate, con gli sporangi raggruppati in
sori, ricoperti o no dall’indusio. Detto indusio può essere fatto a rene, a
cerniera, a scudo; è un carattere diagnostico.
C’è sempre l’anello, che non ha avuto sempre la
stessa posizione, ma ha subito uno slittamento filetico. Infatti, può essere
una linea, tipo l’equatore terrestre, presente in sporangi primitivi, poi
diventa obliquo, poi verticale con passaggi intermedi. Quando è equatoriale, si
ha libertà d’espansione in tutta la circonferenza, quando è verticale è
interrotto nel punto d’inserzione del peduncolo.
Altra cosa interessante è la posizione dei sori,
posti in posizione marginale o nella pagina inferiore. Si ritiene che il tipo
marginale sia più antico, ma in ciascun ordine abbiamo uno slittamento filetico,
per cui troviamo insieme gruppi più primitivi con sori al margine e gruppi più
moderni con sori in superficie. È evidentemente, quest’ultima, una posizione
di vantaggio, perché tutta la fronda è a funzione protettiva.
Per la forma dei sori distinguiamo: sori lineari o
cenosori; sori puntiformi o semplici.
Fondamentalmente due sono i tipi di spore: trilete o
monolete. Nel primo caso è il solito insieme di spore a dare una cellula
tetraedrica a base curva, con tre linee di contatto fra le spore; nel secondo
caso, è la solita “mela tagliata in quattro”, con una sola linea di
contatto fra una spora e l’altra.
I principali ordini sono: Schizeales, a sori
marginali; Cyatheales, cui appartengono le Cyatheae, felci arboree,
formanti imponenti aggruppamenti nel sottobosco; presenti in Etiopia, nella
regione del Caffà; Blechnales, che hanno gli sporangi sulla pagina
inferiore; Polypodiales, con i sori a puntini; Hymenophyllales, a
fronde molto delicate, monostromatiche, molto simili ai muschi e di piccole
dimensioni, misurando 4-5 cm.
Eterosporee e differenziate sessualmente a livello
del gametofito; abbiamo anche macro e micro sporangi diversi, per cui la
differenziazione è spinta e precoce. Sono acquatiche, radicanti e assomigliano
a quadrifogli; sono presenti negli acquitrini.
Hanno prefogliazione circinnata. Ciò che è interessante, sono le profonde trasformazioni delle pinne per dare il sorocarpo. È fatto da due pinne che si chiudono una sull’altra come due valve di conchiglia, saldandosi lungo il bordo, sono anche provviste di un anello gelatinoso lungo la saldatura. Dentro questa conchiglia e attaccati all’anello, stanno i sori, non più aperti, ma completamente chiusi, per cui troviamo i macrosporangi centrali e i microsporangi periferici. Non è completamente un’entità chiusa, ma può avere un orifizio verso l’alto, solo che in genere è otturato. Il nome sorocarpo è comprensivo di quell’entità costituita da due pinne chiuse, ornate di un anello gelatinoso della stessa lunghezza del perimetro delle pinne chiuse. A maturità, quando le pinne si aprono, l’anello stesso si allunga, permettendo l’uscita degli sporocarpi.
Il sorocarpo è completamente chiuso, salvo che per
l’orifizio; la sua parete non è altro che un indusio che si è completamente
chiuso. Le macrospore sono in genere una per macrosporangio, perché delle
quattro entità generate per meiosi, tre degenerano.
I microsporangi producono invece da 32 a 64
microspore.
 Rappresentiamo lo sporofito (1), con parte sterile fatta da quattro
pinne disposte a quadrifoglio e alla base i sorocarpi a diverso grado di
sviluppo. Dentro i sorocarpi (2) ci sono gli sporangi (3) e il tutto è
circondato dall’anello gelatinoso. Al (4) c’è un solo sporocarpo con al
centro quattro macrosporangi e alla periferia i microsporangi. Aprendosi,
lasciano uscire le microspore (7) che germinano per successive moltiplicazioni
cellulari. Nascono, ora, due soli anteridi (11), scarsi nella produzione in
quanto danno solo 16 spermatozoi ciascuno. La macrospora germina all’interno
del macrosporangio e si ha la formazione di un gametofito a funzione trofica
eccetto che per la cellula apicale, che darà un solo archegonio. Avviene la
fecondazione fra lo spermatozoide (12) e l’oosfera, all’interno dell’archegonio
(6) e si forma l’embrione (13), che nutrito dal gametofito, per un certo
periodo, emette una radichetta, l’abbozzo delle foglioline (14) e infine, ridà
lo sporofito adulto. Da ricordare che la differenza sessuale è già a livello
di macro e microsporangio.
Rappresentiamo lo sporofito (1), con parte sterile fatta da quattro
pinne disposte a quadrifoglio e alla base i sorocarpi a diverso grado di
sviluppo. Dentro i sorocarpi (2) ci sono gli sporangi (3) e il tutto è
circondato dall’anello gelatinoso. Al (4) c’è un solo sporocarpo con al
centro quattro macrosporangi e alla periferia i microsporangi. Aprendosi,
lasciano uscire le microspore (7) che germinano per successive moltiplicazioni
cellulari. Nascono, ora, due soli anteridi (11), scarsi nella produzione in
quanto danno solo 16 spermatozoi ciascuno. La macrospora germina all’interno
del macrosporangio e si ha la formazione di un gametofito a funzione trofica
eccetto che per la cellula apicale, che darà un solo archegonio. Avviene la
fecondazione fra lo spermatozoide (12) e l’oosfera, all’interno dell’archegonio
(6) e si forma l’embrione (13), che nutrito dal gametofito, per un certo
periodo, emette una radichetta, l’abbozzo delle foglioline (14) e infine, ridà
lo sporofito adulto. Da ricordare che la differenza sessuale è già a livello
di macro e microsporangio.
Sono piante antiche risalenti all’Eocene (terziario) e giunte sino a noi; sono comuni ed hanno un’estrema velocità di riproduzione. Sono natanti, acquatiche, liberamente galleggianti. Hanno prefogliazione diritta, cosa eccezionale per delle felci e difficile da spiegare, perché non si conoscono gli antenati; la prefogliazione diritta esisteva nelle Ophyoglossidae, ma la parentela è assurda. Le radici della Salvinia sono foglie trasformate. La pianta in origine ha un verticillo di tre foglie, di cui due restano in superficie come tali, mentre la terza, si affonda e si trasforma in radice. Per la riproduzione, non abbiamo sorocarpi, cioè si ha una semplificazione rispetto alle precedenti. Abbiamo solo gli sporocarpi per cui la differenziazione sessuale è ancora più precoce, già a livello dello sporocarpo. L’archegonio delle Marsileidae era unico, qui n’abbiamo diversi per gametofito femminile; nel gametofito delle Marsileidae non abbiamo parti verdi, mentre qui abbiamo una parte a ventaglio verde, su cui stanno gli archegoni. Lo sporocarpo delle Marsileidae ha pareti monostratificate, mentre qui la parete, cioè l’indusio, è doppia, ma tra i due strati di cellule c’è una camera d’aria per il galleggiamento; gli sporocarpi galleggiano, fatto questo importante, perché sono piante natanti. È un adattamento spinto alla vita acquatica. La pianta ha un fusticino piuttosto allungato, verticilli di tre foglie di cui una si trasforma in radice. Produce gli sporocarpi sessuati, distinti in maschio e femmina. Nel primo troviamo numerosi microsporangi con lunghi pedicelli, nel secondo il numero di macrosporangi è ridotto. Anche qui abbiamo una sola macrospora, sempre per il fatto della degenerazione degli altri tre nuclei meiotici. Nel microsporangio, invece, si formano numerose spore. Entrambi i tipi di spore germinano internamente alle pareti dei propri sporangi, perciò dal macrosporangio, sporge in fuori, dalla parete, la lamina a ventaglio, che non è altro che il gametofito femminile con gli archegoni. La parte maschile, germina all’interno del microsporangio e dà due anteridi, a volte uno solo. Il numero di spermatozoi è ridotto, sei nel primo caso, quattro al massimo nel secondo. È un sistema primitivo, in contrasto con il fatto che sono piante evolute più delle felci. Si ha poi fecondazione e lo zigote cresce protetto dalla parete della macrospora e del macrosporangio, fino a raggiunta indipendenza.
Salviniidae e Marsileidae costituivano il gruppo
delle Hydropteryidae, ma si è visto che sono piante molto diverse fra loro,
quindi sono state separate. Alle Marsileidae si fanno appartenere i generi
Marsilea a quattro pinne, Renellidium a due pinne, Pilularia con una sola pinna,
molto ridotta, tutte appartenenti ad una sola famiglia.
Alle Salviniidae appartengono due famiglie, le
Salviniaceae e le Azolaceae, quest’ultime presenti anche da noi, ma americane
come origine, sebbene da noi si siano trovati dei fossili. Si ricordi che
nell’Azolla vive l’Anabaena, una Cyanophyta.