|
Citologia |
La Cellula
|
BIOLOGIA DELLA CELLULA La teoria cellulare. La cellula procariota: generalità, parete cellulare, flagelli, fimbrie, membrana plasmatica, citoplasma, materiale genetico. La cellula eucariota: confronto con quella procariota; la compartimentazione come elemento strutturale e funzionale. Struttura e funzione delle membrane. Il citoscheletro. Il nucleo. Il reticolo endoplasmatico. I lisosomi. L’apparato del Golgi. I mitocondri. Confronto tra cellula animale e vegetale. Riproduzione, aggregazione e differenziamento cellulare. La mitosi. Le strutture tipiche della cellula vegetale. Sistema plastidiale, cloroplasti, cromoplasti, amido. Il sistema vacuolare (inclusioni solide). Struttura, funzione e modificazioni della parete cellulare. Introduzione allo studio del metabolismo. La cellula e l’energia. Il flusso di energia. ATP e lavoro cellulare. Fermentazione, respirazione aerobica ed anaerobica. Gli enzimi. Le principali vie metaboliche e il loro controllo. La fotosintesi C3, C4, CAM. Metabolismo speciale: i cammini biosintetici, le principali vie biosintetiche speciali. |
|
Definizione di Cellula
Nel 1856 LEIDIG definì la cellula come una massa di citoplasma provvista di nucleo. Questa è la più semplice definizione di cellula che ancora oggi si può dare. La cellula è l'unità fondamentale vivente. Generalità La scoperta della cellula spetta al fisico inglese Robert HOOKE (1667) che esaminando una sottile sezione di sughero per provare l'ingrandimento di primi rudimentali microscopi, ne vide la struttura a cellette; ma è solo dopo le numerose osservazioni di Marcello MALPIGHI (1675) che si considerano le cellule (da lui dette otricoli) come costituenti elementari delle piante. MALPIGHI osservò anche le cellule adipose degli animali. Un precursore di questi primi osservatori; fu un commerciante olandese di stoffe, LEEUWENOEK (1632-1723); che, con primitivi microscopi da lui costruiti per contare i fili dei tessuti, potè scoprire e descrivere i globuli rossi; la struttura striata delle fibre muscolari, i movimenti protoplasmatici delle uova di Unio, gli spermatozoi e moltissime altre strutture microscopiche allora del tutto ignote. Nel 1809 BRISSEAU-MIRBEL rimise in auge il nome di cellula che era stata chiamata utricolo per tutto il secolo XVIII. A LEEUWENOEK si deve anche la scoperta del nucleo dei globuli rossi dei pesci; l'esistenza del nucleo fu poi osservata dal FONTANA (1781) nelle cellule epidermiche in Anguilla e da CAVOLINI (1787) nelle cellule uovo dei pesci, ma fu solo nel 1831 che il BROWN potè concludere essere il nucleo un costituente essenziale e costante delle cellule. Nel 1850 von MOHL conclude che "ogni nucleo deriva per divisione di un nucleo precedente".
|
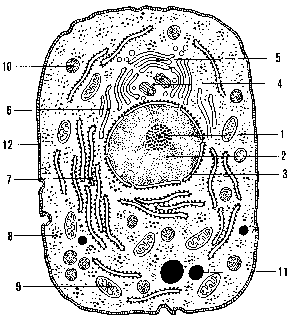
Fig. 1 - Schema cellula ideale
|
La teoria cellulare Nel 1838 SCHLEIDEN per i vegetali, e quindi nel 1839 SCHWANN per gli animali, formularono la teoria cellulare secondo la quale tutti i viventi sono formati da una o più unità viventi elementari: le cellule. Secondo questa teoria i pluricellulari sono da considerarsi come una colonia di organismi monocellulari aggregati insieme. Questo concetto "coloniale" degli organismi pluricellulari ebbe gran fortuna poiché rispecchiava assai bene il concetto evoluzionista che si andava imponendo in quel tempo. Con tale concezione gli organismi più complessi sarebbero derivati, nella storia evolutiva, da cellule che si sono aggregate in colonie via via più complesse. Alcuni Flagellati rappresentano le colonie più semplici; le Spugne rappresentano una condizione coloniale più complessa, dove le cellule non sono tutte eguali in quanto si instaura una divisione del lavoro tra i vari partecipanti alla colonia. Questa differenziazione si va via via esaltando salendo la scala della complessità zoologica. Il concetto della divisione del lavoro, che dobbiamo a MILNE-EDWARDS (1850, è senza dubbio uno dei fondamentali della Biologia e mantiene tuttora tutto il suo valore, come vedremo nei capitoli che seguiranno. Nel 1850, MOHL scopre la divisione cellulare nelle alghe filamentose; NAEGELI nelle piante superiori, e VOGT, KOELLIKER, REMAK, per citare solo alcuni nomi, negli animali. La divisione del nucleo fu osservata soltanto più tardi, nel 1879, da STRASBURGER nelle piante). La teoria cellulare si arricchisce di una nuova conferma, dimostrando l'origine di tutte le cellule di un organismo da una sola cellula iniziale (l'uovo) per divisioni successive. Viene formulato da VIRCHOW l’aforisma "omnis cellula e cellula” e, quale patologo, concepì la teoria che le malattie siano dovute ad alterazioni delle cellule che si manifestano durante la divisione cellulare, ribadendo il concetto che i processi vitali dell'organismo pluricellulare sono dipendenti da quelli delle singole cellule che lo compongono. La teoria cellulare trovò un forte sostenitore in HAECKEL, convinto partigiano della teoria trasformista dell'evoluzione. Egli, anzi schiavo del preconcetto, considerò l’uovo una masserella protoplasmatica priva di nucleo, simile ad ipotetici organismi primordiali da lui chiamati "monere" che con la fecondazione acquisterebbe il nucleo. Ma se l'idea evolutiva portò anche ad aberrazioni concettuali, la sua suggestività ebbe allora il benefico effetto di stimolare le ricerche citologiche ed embriologiche. E' infatti in questo periodo che sorge e si perfeziona la tecnica microscopica. Con la scoperta di granuli nel citoplasma si credette di confermare le antiche concezioni di BRUCKE (1865) e di KOELLIKER (1868) che la cellula a sua volta fosse composta da unità viventi di ordine inferiore. BUFFON chiamò questi granuli "molecole organiche" e HAECKEL "plastiduli”. Ma questa "teoria dei bioplasti" (omnis granulo e granulo) ebbe breve successo poiché le esperienze dimostrarono che non era possibile tenere in vita, isolato dalla cellula» non solo un granulo, ma nemmeno un frammento cellulare (vedi esperienze di merotomia di BALBIANI nei protozoi par.33). Una espressione moderna di questa teoria, che ogni tanto risorge, è la considerazione che nella cellula vi possano essere dei granuli col valore di simbionti, quali sono i cloroplasti, secondo LUYET (1940), e le placchette vitelline delle uova (come aveva già supposto GOLGI e oggi è sostenuto da PIERANTONI e da BUCKNER). Ma questa concezione è tutt’altro che accertata. Critiche alla teoria cellulare Verso la fine dell’800 si cominciarono a formulare le prime critiche alla teoria cellulare. Innanzi tutto si cominciò a criticare la tecnica microscopica e molte strutture, pazientemente descritte nelle cellule fissate e colorate, vennero considerate degli artefatti. La famosa questione se il protoplasma abbia struttura granulare (ALTMANN, MAGGI), filare (FLEMMING), reticolare (FROMANN), o alveolare (BUTSCHLY) viene messa nel giusto valore dimostrando che queste strutture sono il risultato del differente effetto coagulante dei vari liquidi fissatori usati (FISCHEB 1899, SCHWABTZ). Viene criticato il concetto coloniale. La vita del Metazoo non appare come la somma della vita delle cellule che lo compongono, ma essa è una vita superiore. Volfango GOETHE e poi Hans DRIESCH combatterono il concetto coloniale ponendo la individualità superiore al di sopra di quella delle parti. DE BARBY concÌuse dicendo che "non sono le cellule che fanno gli alberi, ma sono gli alberi che fanno le cellule", intendendo dire che l'attività delle cellule è regolata dalla unità superiore, l'albero. La vita dell'insieme e' al di sopra della vita delle singole vite cellulari. Viene infine attaccata l'esistenza della cellula stessa. Il RHODE, all'inizio di questo secolo, fa una Istologia senza considerare le cellule, descrivendo i tessuti quali sincizi nei quali la sostanza intercellulare ha altrettanto valore di quella cellulare. Non vi è dubbio che nelle forme superiori la individualità cellulare è molto spesso compromessa per la presenza di ponti protoplasmatici tra le cellule (negli epiteli, nei connettivi, ecc.); così esistono strutture, quali le fibre muscolari striate, in cui non vi sono cellule distinte, ma, in un'unica massa protoplasmatica si vedono molti nuclei. E molti altri, vedremo, sono gli esempi di sincizi (cellule che si fondono) e di plasmodi (nuclei che si dividono senza la divisione del citoplasma) (fig.7). E' stata cosi' contrapposta alla dottrina cellulare una dottrina "simplastica" (STUDNICKA) ovvero di un protoplasma continuo. Organismi tipicamente acellulari, come alcune alghe (Caulerpa) in cui vi sono numerosi nuclei in un citoplasma indiviso ed altre in cui vi e' un solo nucleo in un organismo piuttosto grande e di struttura complessa (Ace tabular i a) (fig.8), vengono presi come esempi in appoggio di questa concezione; si ottennero sperimentalmente animali senza divisioni in cellule (larve di Chaetopterus: LILLIE). Altro argomento portato contro la teoria cellulare e' quello dei tessuti in cui prevale la sostanza intercellulare con complessità' assai notevole (metaplasmi di HEIDENHAIN), Secondo ASCHOFFimetaplasmi avrebbero una vitalità indipendente dalle cellule. Le recenti ricerche di HUZELLA mettono in chiaro valore queste strutture intercellulari nella organizzazione dei Metazoi. Il concetto di SCHWANN che "... anche di recente da LUYET (1940) in base ai seguenti fatti: a)organismi cenocitici non divisi in cellule: Mixomiceti plasmoidali, i Ficomiceti (alghe brune) ,i Sìfonati (alghe verdi) quali la Caulerpa e la Bryopsis; b) le organizzazioni unicellulari, quali i protozoi e le alghe unicellulari (Acetabularia) che sono ben più di una cellula; e) unità viventi intracellulari quali i cloroplasti (ritenuti da LUYET simbionti); d) la possibilità di vita senza nucleo (esperienze di HABWEY con la centrifugazione); e) le Cianoficee con nucleo diffuso; f) i virus che sono solo nucleoproteine.
Concetto moderno della dottrina cellulare Queste concezioni tuttavia non distruggono il valore di quella individualità delle cellule che alcuni autori non vogliono ammettere. Vi sono semplici osservazioni che dimostrano ciò: alla morte di un organismo non muoiono contemporaneamente le parti; anzi è stato dimostrato che i tessuti muoiono in tempi differenti dalla morte dell'insieme. Si può parlare, paradossalmente, come già fece il TIRELLI (1897), di una "vita del cadavere". Il metodo delle colture in vitro dei tessuti, portato al suo perfezionamento da CARREL, e che oggi si può fare su cellule perfettamente isolate le une dalle altre, permette di dimostrare in modo inoppugnabile questa individualità vitale delle cellule dei metazoi. Si sono potute coltivare cellule di embrione di pollo (fibrobÌasti del cuore) per oltre 20 anni, cioè per un tempo molto più lungo della durata di vita di un pollo. Queste cellule inoltre si sono incessantemente riprodotte e lo sperimentatore è stato costretto ad asportare ogni pochi giorni una parte della cultura. Teoricamente le cellule isolte da un organismo si moltiplicano indefinitamente. Non possiamo pertanto considerare superata la teoria cellulare, ma dobbiamo semplicemente modificarla secondo le nuove acquisizioni. Non vi è dubbio che le cellule derivano da cellule e tutte derivano dalla cellula uovo. Le connessioni tra le cellule (plasmodesmi), la formazione di sincizi, ecc. sono adattamenti che si verificano successivamente, ma anche in un sincizio ogni nucleo domina il proprio citoplasma così che, se non vi è una suddivisione morfologica in cellule, esiste una suddivisione fisiologica. Si è dimostrato come, distruggendo un nucleo di un sincizio con l’ago del micromanipolatore, muore anche una parte del citoplasma, quella sotto il suo dominio. Rimane invece contro la concezione classica, la constatazione che la vita superiore è assai più della vita delle cellule sommate insieme. Possiamo pertanto concludere con COTRONEI (1938) che "... l'organismo è una associazione di elementi dotati di relativa autonomia che costituiscono una, entità di ordine superiore che domina e dirige la vita dei singoli elementi. Questa dipendenza della vita dei singoli elementi dalla vita dell’organismo possiamo ritenerla tanto più accentuata quanto più elevato è il grado di organizzazione. 7 bis. Appendice al paragrafo 7: Colture dei tessuti Metodo delle colture i» "vitro".- Il metodo della osservazione di elementi cellulari sopravviventi è stato impiegato già anni addietro per lo studio del sangue (JOLLY 1898), ma il metodo è diventato una vera "coltura" di cellule vive ad opera di HARRÌSON eh e ha coltivato in linfa di rana frammenti di tessuto nervoso dello stesso animale. Il metodo è stato poi ampiamente perfezionato da altri autori, e specialmente dal CARREL, applicando
8. Colonia e associazione cellulare. Divisione del lavoro. Se il concetto coloniale per i Metazoi è caduto, il concetto di divisione del lavoro è rimasto. Tuttavia possiamo constatare come le prime aggregazioni cellulari comincino a formarsi con carattere di vera colonia. Interessanti esempi ci sono dati dai flagellati: nella Eudorina si ha la condizione di una colonia di 32 cellule simili; colonia temporanea in quanto le 32 cellule si possono separare e divenire altrettanti capostipiti di nuove colonie (fig.); nel Dinobryon la colonia è fissa e gli elementi più vecchi muoiono costituendo una sorta di fusto a supporto della colonia; nel Volvox si ha la prima distinzione di un soma e di una linea germinale, venendo così a rappresentare una condizione di passaggio dalla colonia all'associazione; è un vero organismo pluricellulare con un soma e cellule germinali: di conseguenza è un primo esempio di un soma mortale e di una linea germinale teoricamente immortale. Vedremo tuttavia come la possibilità di riproduzione vegetativa in alcune forme rendono meno assoluta la differenza tra linea germinale e linea somatica. Nei Mesozoi e nei Poriferi assistiamo ad una prima divisione del lavoro tra le cellule del soma con la differenziazione di diversi tipi cellulari. Il passaggio dalla vita coloniale a quella di associazione superiore, che impone un concetto unitario superiore e una vita superiore dell'insieme, si stabilizza con gradualità durante lo sviluppo anche nelle forme multicellulari superiori. Infatti i blastomeri di un uovo in segmentazione possono sovente avere lo stesso valore di una colonia temporanea di Flagellati: possono infatti separarsi e ognuno dare origine ad una nuova individualità superiore (poliembrionia). E' con la divisione del lavoro che, nei Metazoi, partendo da elementi cellulari di eguale aspetto, quali sono i blastomeri, si vanno differenziando i vari tipi cellulari dei vari tessuti, destinati a diverso lavoro funzionale. Per comprendere questo concetto del differenziamento è necessario tener presente come tutte le cellule posseggano le proprietà fondamentali (o primitive) del protoplasma: la eccitabilità, la conduzione degli eccitamenti, la reattività nelle sue varie forme (movimento, secrezione, luce, ecc.), l’attività metabolica con i vari fenomeni collegati. Secondo MINOT, un profondo studioso del problema, "tutte le cellule hanno queste proprietà fondamentali, ma quando una cellula diventa differenziata, benché non perda nessuna delle proprietà fondamentali inerenti alla vita, essa viene a diversificarsi, dalle altre cellule perchè una delle sue proprietà si manifesta in modo più cospicuo: perchè acquisti questa proprietà caratteristica, cioè tale eccesso di sviluppo di una determinata funzione, è necessaria una modificazione della sua struttura". Così col differenziamento, le varie cellule hanno bisogno della collaborazione delle altre per supplire a quelle proprietà che sono rimaste in condizione elementare. La cellula differenziata non ha la possibilità di riprodursi più: è destinata ad invecchiare ed a morire, compiuto che sia il suo ciclo vitale caratteristico, ed è quindi incapace di una vita indipendente. Vedremo nel paragrafo sull'attività riproduttiva dei tessuti (par. 64) come nello stesso organismo i vari tipi cellulari si comportino diversamente rispetto a tale attività e come alcuni tipi cellulari abbiano un ciclo breve, muoiano, e vengono sostituiti da altre cellule giovani, grazie all'attività riproduttiva di elementi indifferenziati del tessuto (tessuti cambiali o meristematici), ed altri vivano quanto vive l'organismo ed abbiano un ciclo vitale corrispondente a quello dell'organismo. E’ stato tuttavia visto che coltivando elementi differenziati in vitro ed aggiungendo alla coltura succhi embrionali contenenti sostanze embrionali attive (i trefoni di CARREL) si ha in genere uno differenziamento ed il riacquisto dell’attività moltiplicativa; solo le cellule nervose non hanno possibilità di differenziamento. Anche il momento, nello sviluppo, del differenziamento è differente per le cellule dei vari tessuti di un Metazoi. |