Sono piante provviste di semi, aplodiplonti,
eteromorfe, con alternanza di gametofito e sporofito. Lo sporofito, predomina
sul gametofito, sia per lo sviluppo sia per struttura e, diversamente che nelle
felci, sempre completamente autonomo dal gametofito.
Il gametofito è ora protetto dallo sporofito, in definitiva si potrebbe considerare parassita il gametofito femminile, che è completamente protetto, mentre il gametofito maschile vive in parte a contatto con l’esterno. Avvenuta la gamia, si origina uno zigote e da questo un embrione che è nutrito dallo sporofito ed è contenuto in uno speciale apparato detto seme che permette la sua conservazione per lungo tempo ed è dotato di sostanze di riserva, necessarie all’embrione durante la sua vita latente. La formazione del seme è caratterizzata dalla disidratazione molto spinta, così da passare da 90-95% d’acqua ad un 10-15%, il che provoca un rallentamento del metabolismo. Il nuovo essere, per riprendere a funzionare del tutto, deve assorbire acqua, ecco perché compare, quando seminiamo un seme, la radice. Da questo momento, siamo di fronte al nuovo sporofito, fatto da fusto, radice, foglie, con la parte sporigena attaccata alla parte aerea e sempre più protetta. È una catena che ci porta infine ad uno sporofito con gli apparati sporigeni chiusi, gametofito chiuso, zigote ed embrione chiusi, fino ad avere sporofito di I generazione, gametofito, sporofito di II generazione uno dentro l’altro, nello stesso individuo. Lo sporofito domina nettamente. Gli apparati sporigeni sono diversi per struttura e sessualità; sono prodotti su rami speciali, ad accrescimento definito; si va sempre più accentuando la riduzione dell’apparato meristematico, fino ad aversi il fiore ermafrodita, terminale, su ramo bloccato.
La differenziazione sessuale è ora a livello delle
foglie. Ecco allora i macrosporofilli trasformati, portanti le parti destinate a
proteggere le spore e gametofito, sottoforma d’ovuli protetti da tegumenti,
che racchiudono una cavità con dentro il macrosporangio e le macrospore, in un
qualcosa che ricorda lo sporocarpo. Il tegumento corrisponde all’indusio;
nelle Angiosperme la parete sarà molto spessa ed è detta nucella.
Gli apparati sporigeni maschili sono fatti in modo da
emettere il loro prodotto all’esterno; si formano delle microspore che si
provvedono di parete spessa e danno il polline che, emesso dal
microsporangio, è portato sulla parte femminile. Si forma, ora, il tubo
pollinico e si ha la vera fecondazione.
È tutto un mondo che va evolvendosi con estrema celerità, attraverso vari tentativi, tutti tendenti all’Angiospermia, cioè alla protezione della parte femminile.
Carattere fondamentale delle Spermatophyta è il
seme, che ha l’importante caratteristica di ibernare, ciò gli permette
d’avere vita latente nel periodo secco. La latenza, specialmente nei luoghi
aridi, può essere di 4-5 anni e la germinazione avviene poi, alle prime piogge
e allora, il ciclo vitale deve compiersi in un brevissimo tempo. Si è
riscontrato che semi poliploidi hanno capacità germinativa molto prolungata.
Le spermatophyta sono divisibili in cinque
sottodivisioni che distinguiamo in base a differenze fisiologiche, morfologiche,
biochimiche. Sono molto antiche, già molto sviluppate nel Devoniano medio,
segno che erano d’origine molto più antica. Difficile è trovare
l’antenato. Da scartare le felci, non solo per la diversa struttura, ma perché
più o meno contemporanee. Probabilmente abbiamo un antenato comune da cui sono
partite le due linee parallele di felci e spermatophyta.
Le spermatophyta, sono poi così diverse fra loro da
non poter avere un unico antenato, sono perciò un gruppo polimetrico;
distinguiamo un progenitore comune per Conifere e Licopsida; un altro per
Filicopsida e le altre spermatophyta. Siamo di fronte ad un sistema monofiletico,
polimetrico. Da criticare la classificazione di comodo in Angiosperme e
Gimnosperme, in base all’esistenza o no di fiori, il che comporterebbe
Conifere e Cicadee insieme; il significato dei due termini è utilitario, non
sistematico. Per gli apparati sporigeni, le possiamo dividere in: fillosporee
se sono portati su entità che sono foglie trasformate e in stachiosporee
se gli ovuli sono portati su uno strobilo (conifere). Ci sono però, piante con
ambedue le caratteristiche.
Importante è l’Angiospermia, cioè il
tentativo di proteggere in vari modi, seguendo diversi modelli di diverso
successo, gli ovuli e sottrarsi all’ambiente esterno; in base a ciò, erano un
tempo distinte in: Gimnosperme, ad ovuli nudi e Angiosperme, ad ovuli racchiusi
nell’ovario, ma oggi questa netta distinzione è superata e non ha alcun
valore sistematico.
Sono piante fossili e così definite in quanto hanno le foglie simili alle felci. Hanno apparati sporofillici senza foglie perianziali proprie, cioè senza calice e corolla, per cui gli ovuli sono a contatto con l’esterno. Il legno secondario è omossilo, fatto quindi dalle sole primitive tracheidi con parete terminale a becco di flauto e perforate.
Sono macrosporofillata, con macrosporofilli pennati e non riuniti in strobili, ma con asse centrale e le foglie laterali a disposizione pennata, il tutto direttamente portato dal fusto. Le foglie sono grandi e si ritiene che la fecondazione avvenisse per mezzo di spermatozoi cigliati, ma essendo fossili, nulla si sa con precisione.
Le due classi in cui vengono suddivise sono affini ma abbastanza diverse.
Le Pteridospermopsida, con struttura più primitiva, dal Devoniano medio, giungono fino al Triassico; le Caytoniopsida, dal Triassico arrivano fino al Giurassico, quasi come se derivassero le une dalle altre.
Sono fra i primi fossili trovati e perciò noti da lungo tempo. Dapprima furono note le grandi foglie, tipo felci, poi si trovarono anche dei semi che si ritennero appartenenti ad entità diverse e furono considerati come Angiosperme. Perfezionandosi gli studi, si vide che i semi avevano delle ghiandole sulla cupola e sulle foglie. Poi furono trovate foglie con i semi attaccati e allora si fu certi di trattarsi di una sola pianta, chiamata dapprima Cycadophylicea, per avere le foglie una cuticola simile a quella delle Cycas e foglie simili a quelle delle felci.
Il fusto aveva protoxilema mesarco, carattere arcaico, accrescimento secondario ben evidente e tracheidi con caratteristiche punteggiature areolate nella parete radiale. Le foglie erano grandi a prefogliazione circinnata, molto suddivise, con spessa cuticola di tipo spermatophytico.
I microsporofilli erano
caratteristici in alcuni gruppi; i microsporangi erano, infatti, raggruppati o
liberi; talvolta formavano sinangi privi d’apertura, cioè sempre exanulati. I
sinangi erano spesso di forma strana, in un genere a forma di bicchiere, con gli
sporangi tubuliformi saldati fra loro a dare una sorta di calice contenente le
spore. Talvolta il “bicchiere” si chiude in cima; è un sistema strano solo
in apparenza, perché essendo rivolti in basso si aveva con facilità la
dispersione delle spore.
I macrosporofilli non erano molto modificati, portavano ovuli con una cupola alla base, intera o frastagliata, con ghiandole uguali a quelle che c’erano sulle foglie. Nell’ovulo o giovane seme, era possibile distinguere tre tegumenti o “testa”, detti sarcotesta, sclerotesta ed endotesta. Questa notevole difesa era probabilmente in relazione ad un’ibernazione. Nel punto d’incontro dei tre tegumenti s’individuava una camera pollinica in cui penetrava il granulo pollinico, attraverso un’apertura detta micropilo.
Dentro i tegumenti si ha la parete dello sporangio, dentro le spore. Si ritiene che la fecondazione, avvenisse sulla pianta, ma poteva forse avvenire quando l’entità era staccata, sebbene fosse più probabile la prima ipotesi, per la presenza della camera pollinica.
Per il significato ed omologia dell’ovulo, immaginiamoci una foglia di Robinia pseudo-acacia. Si ritiene che la cupola e i tegumenti derivassero da una foglia pennata, dove la foglia apicale era l’ovulo, per riduzione progressiva della foglia stessa. Avevamo così una foglia impari pennata, dove la pinna terminale era l’ovulo. Ora per accorciamento progressivo dei tratti di rachide fra le pinne laterali, si aveva una loro conversione verso la base dell’ovulo. Per saldatura attorno all’ovulo, si aveva la cupola.
Ricordiamo i generi Lyginopteris oldamia, nota come tronco, mentre i suoi semi si dicevano appartenere a Lagenostoma lomaxi, fino a quando non furono trovati semi e tronchi insieme; della Neropteris heterophylla, si trovò che dal tronco partivano le fronde e alla loro base c’erano i semi.; così Oliver Scott, nel 1903 aveva trovato tronco e semi; nel 1904 a Kingston si trovarono foglie e semi, e fu possibile, in tal modo, ricostruire la pianta.
Risalgono
al Triassico e Giurassico. Avevano foglie rettinervie, pennate o palmate,
quindi, come modello, erano abbastanza evolute. I microsporofilli, erano portati
da pedicelli in posizione apicale a gruppi di cinque o sei. Ogni microsporofillo
è un sinangio di quattro entità (tetrapteri) e tetraloculari. Il polline era
fatto da una massa centrale e da due sacche laterali vuote, aerifere, per
aumentare il galleggiamento, e che ritroveremo nelle Conifere. I macrosporofilli
erano formati da tante pinne fertili, esse si piegavano al margine e si
saldavano; si aveva così un’entità chiusa a parte di un piccolo forellino
basale. Sulla pagina interna della pinna, si trovavano gli ovuli, protetti. Ciò
è valida per ciascuna pinna, per cui si aveva un insieme d’entità fertili.
Gli ovuli erano ben differenziati, il granello pollinico poteva entrare
attraverso il micropilo e germinare (impollinazione micropilare). È un’angiospermia
parziale perché la cavità non è completamente chiusa; inoltre non è la
foglia tutta che si racchiude, come nelle piante superiori, ma sono le singole
pinne. È un modello che sarà poi, abbandonato.
Quando le Caytoniopsida furono scoperte, si ritennero antenate delle Angiosperme, ma si vide subito che l’angiospermia era parziale e la cavità non era chiusa; che non si aveva un ovario, perché non era tutta la fronda a ripiegarsi, ma solo una pinna; che l’impollinazione era ancora micropilare, direttamente sull’ovulo e non stigmatica, come sarà nelle Angiosperme. È però un gruppo importante perché ci dice che già da ora, si ha tendenza a racchiudere gli ovuli in una cavità.
Sono affini alle Pteridospermopsida, ma è difficile
trovare loro discendenti, dato che siamo ancora di fronte a sinangi;
modernamente si è messo in evidenza che questi singoli tetrapteri e
tetraloculari ricordano un po’ le antere, per cui sarebbe considerabile come
modello per la realizzazione di detta entità.
Sono quasi tutte piante fossili, con soli nove generi e presenti in Italia con la sola Cycas d’origine orientale. È una linea conservatrice che ha avuto enorme sviluppo nel passato e che va pian piano scomparendo. Le specie tropicali sono difficili da coltivarsi, più facili gli esemplari orientali.
Il legno secondario è omoxilo, di sole tracheidi. Le foglie sono grandi, estese.
Caratteristica è, al centro della corona di foglie,
la corona di fronde fertili, che nascono alternativamente, un anno si ed uno no.
La parte fertile della fronda è quella basale,
quindi è qualcosa di molto diverso dalle Pteridospermophytina, dove l’ovulo
era la pinna centrale apicale e le foglie basali erano a protezione con la
cupola; qui la parte alta è difensiva, la basale fertile. Spesso la parte
sterile si è trasformata e saldata in squame e porta alla base due ovuli.
Hanno apparati sporofillici senza foglie perianziali
proprie. I macrosporofilli sono in gruppi o in strobili di tante squame saldate
insieme, o anche soli gruppi di fronde a corona all’apice del fusto.
La fecondazione avviene per mezzo di spermatozoi
cigliati, che non nuotano più in un liquido esterno, ma in quello prodotto
dalla pianta nella camera archegoniale
Sono piante fossili del Triassico, ma giungono fino a
noi. I fossili sono ben conservati, specialmente le foglie, per cui ci è
possibile vedere come non siano molto trasformate; è perciò una linea
conservatrice.
Sono piante dioiche, con individui maschili separati
dai femminili; i maschili, da noi, sono piuttosto rari. Il fusto è colonnare,
talvolta di soli 15-20 cm., altre volte fino a 1-2 m. All’apice si trovano le
foglie, sterili, grandi, espanse, consistenti perché abbondantemente
cutinizzate; per l’anatomia, troviamo, meristemi apicali definiti o no, il che
comporta la comparsa di due grandi gruppi a fusto ramificato monopodiale o
simpodiale; nel I caso il meristema è indefinito e ogni anno si accresce senza
soste, non solo in altezza, ma dando anche una nuova corona di foglie o di
fronde alternativamente, anzi si ha dapprima la produzione di una corona di
squame appuntite all’apice, corte, ridotte; l’anno dopo, dà la corona di
foglie, l’anno dopo ancora, la corona di foglie fertili, poi il ciclo
ricomincia.
Il fusto a ramificazione simpodiale è in relazione
al meristema bloccato, producente all’apice strobili maschili o femminili e
poi cessa di funzionare. La funzione accrescitiva, è ora assunta da brevissime
ramificazioni laterali, pressoché invisibili, da cui il nome di simpodiale,
dato che sono i rami laterali ad accrescersi.
Le prime corrispondono alla famiglia delle Cycadaceae,
le seconde alla Zamiaceae.
Il fusto giovane è un’eustele, con diversi fasci
circolari, collaterali, equidistanti, con floema e xilema sullo stesso raggio,
protoxilema endarco. Il midollo è grande, abbiamo canali mucipari danti
mucillagini abbondanti. L’accrescimento secondario comporta un anello continuo
di xilema.
Interessante è il caso delle radici; sono molto
ridotte, perché fin da giovanissime, attaccate da batteri che ne provocano la
necrosi. Nelle zone necrotiche s’impianta la Anabaena cycadearum, una
cyanophyta. È un’alga azotofissatrice, formante degli agglomerati notevoli
che danno alla radice il singolare aspetto coralloide. Queste radici sono
inoltre superficiali, perché devono fissare azoto atmosferico. È una simbiosi
importantissima. L’apparato radicale ridotto, comporta spesso l’abbattimento
per opera del vento, per questo motivo, dette piante si difendono non
raggiungendo mai altezze eccessive.
L’accrescimento è molto lento, probabilmente per
il ridotto assorbimento di sostanze nutritizie.
La prefogliazione è parzialmente circinnata; la
rachide ha, infatti, prefogliazione diritta e sono le pinne ad averla circinnata.
Sezionando lo stipite, abbiamo due fasci a
protoxilema mesarco; si spiega ciò ammettendo che nel passaggio alla struttura
secondaria si è avuto un incontro dei due protoxilema, per cui è qualcosa di
simile ad una stele anfifloica.
Mentre nelle felci avevamo sori, sporangi e spore
libere nel terreno dove germinavano a dare il gametofito, qui le spore germinano
sul macrosporofillo, qui si sviluppa pure il gametofito, qui si ha la
fecondazione, qui si hanno i semi; insomma tutto è attaccato alla fronda. Solo
il seme maturo si stacca e mentre prima potevamo considerare i macrosporofilli
indipendenti dall’apparato riproduttore, qui anche gli apparati riproduttori
sono sul macrosporofillo.
Come già detto, i macrosporofilli sono a corona,
all’apice del fusto o in strobili, grandi anche 50-60 cm. Gli strobili sono
fatti da un asse centrale attorno al quale sono i macrosporofilli, formati da un
supporto centrale detto stipite, ovuli basali, parte apicale fusa che
protegge gli ovuli fino a maturità, quando va in disfacimento; ritroviamo anche
qui, la parte sterile e la fertile che avevamo visto nelle fronde.
Ogni ovulo è ortotropo, nonostante le apparenze
contrarie; è provvisto di un tegumento ben vascolarizzato, dentro cui si trova
il macrosporangio o nucella, voluminoso, massiccio, a parete non
monostromatica, come invece era nelle felci. I tegumenti rappresentano l’indusio
trasformato, la nucella è nome indicante anche la parete del macrosporangio
contenente le spore. La parete del macrosporangio è saldata ai tegumenti,
eccetto che nella parte apicale, dove c’è una camera pollinica, nel cui
interno, troviamo le cellule madri delle spore, danti quattro macrospore
impilate, a diverso sviluppo, di cui tre degenerano. Ne resta una sola, la più
grossa e la più interna, che germinerà e darà il gametofito femminile.
Sviluppandosi più come parte trofica, che come parte fertile, lascia sotto la
camera pollinica, dentro la parete del macrosporangio, una camera archegoniale.
Abbiamo quindi un tegumento con micropilo, apertura su cui giunge il granello di
polline che penetra nella camera pollinica sottostante; questa è immediatamente
seguita e quindi separata dalla parete del macrosporangio, cui fa seguito la
camera archegoniale, che è la parte fertile e trofica di quell’entità che è
il gametofito femminile che ha occupato, sviluppandosi, tutta la parte interna
del macrosporangio.
Il gametofito è ora formato da una parte trofica
voluminosa, dapprima cenocitica e poi cellularizzata detta endosperma primario,
verso l’alto troviamo, infossati nell’endosperma, due archegoni, fatti
ciascuno da una grossa oosfera, con piccoli residui costituiti da una cellula
del ventre e due del canale. Quindi, l’oosfera è infossata nell’endosperma,
protetta dalla parete del macrosporangio, da cui è separata dalla camera
archegoniale, che successivamente, si riempirà del liquido in cui nuoteranno
gli spermatozoi, e dai tegumenti più esterni. L’oosfera è grossa circa ½
mm. per cui è visibile ad occhio nudo.
I microsporofilli sono squamiformi o peltati, ultima trasformazione della squama. Sono riuniti in strobili, costituiti da tante squame, sotto ognuna delle quali ci sono i microsporangi o sacche polliniche. Solo da questo momento è lecito parlare di nucella e di sacche polliniche, non per i gruppi inferiori. Dette sacche sono poste al lato dorsale della squama, cioè verso il terreno, per cui le spore escono facilmente, quando questa si apre. Le microspore, si possono già definire come granulo pollinico. Presentano un notevole indurimento della parete esterna, ulteriormente protetta da strati di sporopollenina. A maturazione, la microspora si stacca, ma è incerto se si stacchi come tale, cioè ancora uninucleato, o se si siano già avute moltiplicazioni del nucleo, per cui potrebbe essere già detto gametofito femminile. Staccatosi, è trasportato sulla parte femminile, processo detto d’impollinazione, cioè, un semplice trasporto e ben distinto dalla fecondazione. Per questo processo d’impollinazione, molte microspore vengono sprecate, ma qualcuna arriva sul micropilo, ma sempre a contatto con l’esterno. Sul micropilo si trovano sostanze che attirano il granulo nella camera pollinica sottostante. Iniziano ora, le trasformazioni e si ha, dentro la parete del granello, la formazione di due cellule protallari rudimentali, residui della parte trofica, ben presto affiancate da una cellula vegetativa e da una cellula anteridiale, la prima sottintendente alla parte trofica in seguito a regressione delle protallari; la seconda destinata a produrre i gameti maschili, cioè a funzione germinativa.
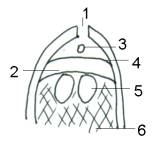
1- micropilo 2
– camera archegoniale 3 –
granulo pollinico
4 – nucella
5 – oosfera 6
– endosperma
È la parte trofica che prolifera per prima, rompendo
la parete del granello e formante il tubetto pollinico, più o meno flessuoso,
con pareti sottili, detto anche budello pollinico; formatosi, esso va ad
accollarsi alla parete interna dei tegumenti, spinge, facendo forza, fino a
forare la parete del macrosporangio ed entra nella camera archegoniale. È un
sistema propulsore a spinta dal dietro. Durante questo processo di spinta, la
cellula vegetativa ha provveduto ad accrescere il tubetto, mentre l’anteridiale
ha dato una cellula parietale che va in disfacimento, poi una cellula madre
degli spermi, che sembra però formarsi dentro la camera stessa, per cui avremo
durante la spinta, solo attività vegetativa. Dentro la cellula madre si ha una
cariocinesi, che porterà alla formazione di due grossi spermatozoi, con una
fila evidente di ciglia, elicoidalmente disposte; sono spermatozoi nudi, cioè
senza parete cellulare e sono i più grossi spermatozoi che si conoscano,
misurando circa 300 micron.
Quando sono maturi, il granello si sfascia, gli
spermatozoi sono liberati dentro la camera archegoniale, nuotano liberamente nel
liquido interno e dirigendosi verso l’archegonio vi penetrano e fecondano l’oosfera,
che è infossata nell’endosperma. In genere si ha la fecondazione di una sola
oosfera, se di due, in seguito una degenera e l’embrione è sempre unico. Il
gametofito maschile è in definitiva ridotto al tubetto pollinico con le due
cellule protallari regredenti, più la cellula vegetativa, più l’anteridiale
che darà la cellula madre degli spermi, più la cellula parietale rudimentale.
Inizia qui la riduzione del gametofito maschile, che poi si ridurrà a soli tre
nuclei.
La fecondazione è qui ancora operata da spermatozoi,
per cui si ha apporto al nuovo essere anche di una parte citoplasmatica
maschile, fatto importante se è vera la trasmissione di certi caratteri ad
opera del citoplasma. Si ha, cioè una fecondazione di nucleo e citoplasma,
mentre nelle piante superiori, avvenendo la fecondazione solo per mezzo dei
nuclei spermatici, solo la madre sarebbe responsabile di certi caratteri
citoplasmatici.
La differenza con le felci, in cui si avevano
spermatozoi nuotanti, è che qui la “nuotata”, avviene in un liquido
interno, prodotto dalla pianta stessa nella camera archegoniale.
Nel corso del tempo, la camera archegoniale scompare,
allorché insorge la fecondazione per nuclei spermatici. Fino a quando ci sarà
la camera, il budello pollinico resta di tipo propulsore, spingendo la parte
basale del granello, poi, diventerà vettore, cioè si ribalterà la sua
posizione, il granello resterà fuori e attraverso il tubo, andranno sulla parte
femminile solo i nuclei.
Per le affinità, sono un gruppo singolare, perché
macrofillata, ma con strobili, carattere questo più proprio delle microfillata,
mentre le macrofillata portano le parti fertili sopra o sotto la foglia. Sono
considerabili allora quale gruppo sintetico in grado di dare origine alle
Conifere da un lato e alle Angiosperme dall’altro, in teoria, almeno. In
pratica la loro discendenza e molto incerta; è un gruppo quindi di notevoli
possibilità evolutive, solo che non le ha sfruttate, è rimasto primitivo, con
scarsa progenitura.
L’ordine delle Cycadales è rappresentato da piante
viventi, con pochi generi; i macrosporofilli sono pennati con 2-8 ovuli
marginali, le foglie sono pennato-sette con un’unica articolazione. Nella
famiglia delle Cycadeaceae, i macrosporofilli sono pennati, con alternata
formazione di una corona di foglie sterili e una di fertili, a spirale, con
meristema indefinito, attorno alla cima del fusto. Comprende il solo genere
Cycas, pianta gerontogea, cioè dell’Eurasia e Africa.
Le Zamiaceae hanno macrosporofilli squamiformi con
due ovuli basali, in strobili, con accrescimento definito per lo strobilo
terminale; alla base dello strobilo insorge una gemma laterale di notevole
vigore in grado di spostare lateralmente lo strobilo, per cui il fusto è sempre
diritto, anche se l’accrescimento è dato da gemme laterali (simpodiale).
Ricordiamo i generi Zamia, americana; Encephalartos,
africana, di 3-4 m. d’altezza e tronchi di 25-30 cm. di diametro; Macrozamia,
australiana, la sola specie presente in quel continente.
Le Nilssoniales, fossili estinte dal Cretaceo
superiore, per cui già quando le Angiosperme sono ben sviluppate. I
macrosporofilli sono peltati, cioè con una parte fertile centrale, al centro di
una squama circolare, da cui pendono due soli ovuli.
Sono piante fossili, poco conosciute. Devono il loro
nome alla costituzione della stele, a cinque fasci legnosi. Sono comuni nei
terreni giurassici dell’India e solo in questa regione sono stati trovati, per
cui hanno avuto sviluppo limitato in tutti i sensi. Il fusto è una stele molto
singolare in cui abbiamo cinque elementi xilematici, più o meno triangolari e
fatti così da avere protoxilema verso l’esterno, ma mesarco. È una posizione
strana e si pensa che avessero protoxilema endarco che per ulteriore sviluppo
della parte xilematica sia stato spostato verso l’esterno. Sarebbe perciò un
prodotto secondario. È una caratteristica che ne permette l’immediato
riconoscimento.
Hanno foglie abbastanza lunghe, intere, inserite su
brachiblasti lineari (macroblasti sono i rami lunghi, brachiblasti quelli corti,
prodotti dalle Conifere, su cui s’inseriscono le foglie).
Hanno strobili più o meno simili alle coccole dei
cipressi, brevi, ovoidali, portati in gruppo, per cui si ha come
un’infiorescenza di strobili.
Gli ovuli sono sessili, addensati insieme; non è
stata evidenziata la camera pollinica, forse perché sono fossili o molto più
semplicemente, perché non c’era.
Il fascio vascolare si prolunga fino alla base
dell’ovulo, forse per il motivo che il seme aveva una parte carnosa esterna.
In definitiva è un gruppo strano, con fusto molto
sui generis.
Hanno stomi simili alle Bennettitopsida; gli
strobili, anche se in infiorescenze, li accomunano alle Cycadophytina. Non sono
considerabili quale gruppo a sé; restano da spiegare varie cose, come la
mancanza della camera pollinica. Dove andava a finire il polline? Dove si
sviluppava il gametofito? Sono fossili esclusivamente indiani.
Gruppo molto importante, ma solo fossile. Appaiono
nel Triassico e arrivano fino al Cretaceo. Hanno fusto colonnare od ovoidale.
Portavano all’apice una corona di fronde; nel suo interno era un midollo
ampio, cosa comune d’altre Cycadophytina. Le foglie sono pennato-sette o
indivise, mai foglie pennato-composte, ma sempre semplici, anche se sempre
profondamente suddivise. Carattere nettamente distintivo, è che sono monoiche,
cioè con i due sessi sullo stesso individuo.
I macro e i microsporofilli, diversi dalle foglie
normali, erano disposti a spirale all’apice del tronco. Ambedue s’inserivano
come ramificazioni laterali, su un asse centrale. I microsporofilli erano
inseriti su una sorta di fronda pennata, con le sacche polliniche inserite sugli
assi secondari, da giovani erano revoluti e solo da vecchi s’aprivano a
maturazione raggiunta. Le sacche erano in numero variabile. All’interno della
corona di fronde, spesso circondate da una serie di brattee ligulate, troviamo i
macrosporofilli, spiralati e formanti uno strobilo. Nel loro insieme le parti
femminili davano un complesso, in cui le varie parti s’inserivano a spirale su
un asse centrale, da cui partivano peduncoli portanti l’ovulo provvisto di un
micropilo molto allungato. Interposte, vi erano brattee sterili interseminali.
Era un insieme chiuso, tondeggiante, per il toccarsi delle brattee, da cui
sporgevano i micropili. Viste dall’alto, le brattee interseminali, erano
rotonde, a stretto contatto fra loro; nel punto d’incontro di tre di esse,
c’era uno spazio libero da cui usciva il micropilo.

È una sorta d’angiospermia, perché l’ovulo era
protetto; avendo la colonna micropilare sporgente, si realizzava
l’impollinazione senza che gli ovuli venissero mai a contatto con l’ambiente
esterno, diversamente da quanto avveniva nelle Caytoniopsida; anche qui è un’angiospermia
incompleta; è un modello che ha avuto successo, ma solo in quel periodo, mentre
è quello delle Caytoniopsida quello da cui deriva l’angiospermia moderna.
Hanno avuto successo ed è il gruppo più avanzato
della sottodivisione, ma non hanno dato discendenti; sono una linea a fondo
cieco.
Le Cycadophytina, sono in definitiva anch’esse una
linea a fondo cieco, che ha dato una linea che è giunta fino a noi con le Cycas,
le Zamie, e altre, e l’altra, le Bennettitopsida, che hanno avuto un successo
notevole, ma si sono estinte, pur essendo abbastanza evolute, forse per essere
arrivate al limite delle loro possibilità.
Le possiamo interpretare quale gruppo sintetico: le
parti fertili sono sia ad inserzione apicale, direttamente sul fusto, sia in
strobili apicali, per cui abbiamo i due tipi, fillo- e stachiosporee. Sono
piante monoiche e dioiche, sono quindi, un poco, la sintesi di Conifere e
Angiosperme, ma malgrado ciò, non ci sono discendenti. Questo fatto singolare
è da mettersi in relazione col modo di vita, cioè col debole apparato radicale
che rende difficile, non solo il raggiungimento di una notevole altezza, ma lo
stesso approvvigionarsi delle sostanze necessarie.
È un nome collettivo per indicare Cordaitopsida,
Ginkgopsida, Taxopsida, conifere vere e proprie, un insieme, cioè, di piante
molto diverse, ma strettamente affini. Le Ginkgopsida sono le ultime con
fecondazione per opera di spermatozoi cigliati.
È un gruppo che ha avuto un vastissimo successo e
popola vaste regioni del globo terrestre, formando foreste di vario tipo,
pinete, abetine, lariceti, ecc. in tutti e due gli emisferi.
Hanno apparati sporofillici senza foglie perianziali
proprie. Il legno secondario è omoxilo di sole tracheidi, per cui sono
abbastanza basse nel livello evolutivo. Non sempre i macrosporofilli sono
portati in coni, ma spesso anche in modo da costituire un insieme biforcato o
pluriforcato come nel Ginkgo, oppure l’ovulo è isolato e provvisto d’arillo
carnoso come nel Taxus.
Le foglie sono sempre di modeste dimensioni e
rientrano fra le microfillata. Una certa eccezione è rappresentata dal Ginkgo,
che essendo un fossile vivente, ha le foglie più grandi, come appunto gli
esemplari fossili del gruppo stesso.
Per la fecondazione, nulla si sa per le Cordaitopsida;
per le Ginkgopsida c’è ancora un certo apporto di citoplasma maschile,
avvenendo per spermatozoi; per le Conifere si ha solo apporto del nucleo
maschile. Il tubetto pollinico è ancora propulsore nel Ginkgo, poi diventa
vettore. Sono queste le principali differenza fra i gruppi.
Sono da considerarsi il primo gruppo, non solo perché
le meno evolute, ma perché gruppo sintetico che riunisce in sé certe
caratteristiche degli altri gruppi e avente per questo una notevole potenzialità
evolutiva.
Compaiono nel Devoniano medio e all’inizio del
Mesozoico si estinguono, pur avendo avuto un notevole successo. Non sono molto
comuni come fossili, ma sono comunque ben conservate.
Erano grosse piante, colonnari, alte 20 m. e
costituenti vere foreste imponenti. Presentando poi, un ampio midollo, si pensa
che vivessero in un ambiente favorevole.
Le foglie, erano in spirale, parallelinervie, senza
costa centrale, non sappiamo se terminassero con forma tonda o a punta, avevano
però i margini paralleli e alla base, erano dicotomicamente divise.
È un sistema che si ritroverà nel Ginkgo, con la differenza che qui il
sistema dicotomico si slarga a ventaglio. Non si ha mai una nervatura centrale.
Gli strobili maschili e femminili, sono formati da
poche entità, nel maschile c’erano brattee esterne sterili, in zona centrale
apicale c’erano i microsporofilli. Le brattee erano fatte in modo simile alle
foglie, con nervature parallele, terminanti a coda di rondine, con costa
centrale; nella biforcazione s’inserivano i microsporangi. Per ogni strobilo
avevamo diversi sporofilli. Il modo particolare della loro struttura, secondo
alcuni, è in discordanza con quella della foglia, perché qui, in definitiva
esiste una costa centrale ben vascolarizzata e provvedente a nutrire i
microsporangi. Si pensa che siano entità interpretabili come un asse centrale,
portante all’apice i microsporangi e resasi alata in un secondo tempo. Questa
teoria è giustificata dal fatto che i granelli pollinici debbono essere
dispersi e le ali, contro cui il vento urta, servono ottimamente allo scopo.
Anche la parte femminile, è organizzata in strobili
e provvista di brattee sterili con l’asse dello strobilo che porta da uno a
quattro macrosporofilli con all’apice uno o due ovuli. Ciascun macrosporofillo
è costituito da un lungo peduncolo con all’apice gli ovuli e una piccola
appendice lungo il peduncolo stesso, appendice interpretata come residuo di una
parte fogliacea; cinque o sei di queste entità costituivano uno strobilo. Si ha
una sproporzione notevole fra il picciolo e la parte fogliacea appendicolare; il
fatto che qui è ridotta, sarebbe conferma che nel maschio si ha parte assile
con ali laterali, non una foglia particolare; in definitiva sia in maschi che
femmine si avrebbe la scomparsa più o meno spinta della parte fogliare, in
relazione ad una maggiore facilità di dispersione, come sembrano confermare
anche le microspore provviste di più sacche periferiche vuote per il
galleggiamento nell’aria.
Il lungo peduncolo, cui stanno attaccati gli ovuli pendenti, si ritroverà nelle Ginkgopsida, dove gli ovuli sono ortotropi, mentre nelle Taxopsida si ha un solo ovulo isolato, come spesso anche qui.
Diverse sono state le possibilità evolutive del gruppo, ma è difficile dire quali Conifere siano da esso discese direttamente. E un gruppo sintetico, di piante molto evolute, derivate da un antenato comune anche alle Conifere.
Unico antenato comune possibile sembrano essere le
Licopsida, dove c’è il sistema dicotomico, gli ovuli solitari, peduncolati.
Le Licopsida fossili, erano poi piante colonnari molto grandi. D’altra parte
non hanno niente in comune con Pteridospermophytina e Cycadophytina, perché
sono macrofillata, mentre qui le foglie sono sempre piccole, se non altro al
confronto con quelle.
È un nuovo modello evolutivo, scomparso, ma che ha
avuto successo per certi sistemi organizzativi.
È un
gruppo che avuto molto successo, ma oggi una sola specie è sopravvissuta, la
Ginkgo biloba, spontanea in Asia e coltivata da noi. È dioica e fatto strano,
è che nell’orto botanico dell’Università di Genova, gli esemplari sono
tutti maschili, i femminili non attecchiscono, mentre a Firenze succede il
contrario. Risalgono al Permiano inferiore e giungono fino a noi. Il fusto è
elevato, piuttosto grande, con esemplari di 300-400 anni. Hanno macroblasti e
brachiblasti, rametti più piccoli su cui sono inserite le foglie. Le foglie
sono disposte a spirale sui piccoli rametti, differenziati e sono o nastriforme
o flagellate, senza costa centrale. Nel Ginkgo a foglie flagellate, abbiamo le
dicotomie basali che poi divergono a ventaglio; nelle foglie nastriformi dei
fossili le nervature sono parallele. Le foglie possono essere sessili o
picciolate; il picciolo è ben sviluppato nel Ginkgo, nei fossili no. Il midollo
è piccolo. Le gemme sono singolarmente provviste di perule che finiscono poi
per cadere.
Le piante sono dioiche, ma ci vogliono circa 20 anni
per vedere che sesso abbia l’individuo nato dal seme. I maschi portano
strobili amentiformi, pendenti, ciascuno fatto da un numero alto di
microsporofilli. Ogni microsporofillo ha una parte di sostegno, due
microsporangi o più, ovoidali, sormontati da un proseguimento di forma
triangolare. È qualcosa di simile all’antera, solo che nell’antera manca
questa sorta d’appendice apicale, residuo del microsporofillo. È interessante
perché ci fa vedere come uno stame non sia altro che un microsporofillo
trasformato. Nei microsporangi o sacche polliniche, si producevano i granuli
pollinici, che erano privi di sacche, fatto in accordo con le Taxopsida,
diversamente che per gli altri due gruppi. I granuli pollinici vanno a
depositarsi sul micropilo, penetrano nella camera pollinica, proseguono nel loro
sviluppo, nel modo visto anche per le Cycas. Il tubetto pollinico s’impianta
nella camera pollinica, spinge il pezzo basale dentro la camera archegoniale, a
volte sfrangiandosi in più frange e prendendo contatto con la nucella per fini
nutrizionali. Arrivato nella camera archegoniale, si sviluppano, al solito, gli
spermatozoi cigliati. L’ovulo corrisponde in pieno a quello delle Cycas, anche
per struttura. La parte femminile, comprende macrosprofilli, fatti da un insieme
completamente fertile, salvo il cercine. Sono provvisti di un supporto più o
meno lungo, bi o pluri-forcato all’apice, portante due ovuli ortotropi. Detti
ovuli presentano in basso un cercine anulare e sopra, ancora parte fertile. La
parte basale è provvista di due cordoni vascolari che vascolarizzano
separatamente i due ovuli. Si è discusso sul significato del cercine che si
pensava residuo fogliare del macrosporofillo, analogo al dente delle
Cordaitopsida. Si è visto non essere possibile sostenere questa tesi, perché
è una caratteristica mancante in certi fossili e anche nella Trichophyta, la più
antica delle Ginkgopsida. È perciò una struttura sviluppata successivamente.
L’ovulo apicale, è fatto dall’esterno, dai
tegumenti, camera pollinica, cellule archesporiali danti la cellula madre delle
spore, in genere unica, che darà poi le quattro spore, di cui tre degenerano e
una sola si sviluppa a dare il gametofito femminile, costituito da una parte
trofica o endosperma primario e da una o due grosse oosfere, provviste dei
residui dell’archegonio, cioè le due cellule del collo e quella del ventre,
come nelle Cycas. La fecondazione interessa ambedue le oosfere, ma l’embrione
è sempre uno. Il Ginkgo si distingue nettamente dalle Conifere per non avere
assolutamente strobili, dato che i microsporofilli sono in amenti, cioè
organizzati in unità pendenti e non erette; i macrosporofilli sono isolati, ma
l’essere a corona ce li potrebbe far considerare, come residui di uno strobilo
ridottissimo. Anche nelle Taxopsida, gli ovuli sono isolati, senza strobilo vero
e proprio; nel Ginkgo, quindi, troviamo dei caratteri che lo accomunano ad altri
gruppi della sottodivisione.
Sono una classe di piante, che non solo ha
abbondantemente colonizzato il mondo antico, ma abbondantemente popola il
moderno; le troviamo nei due emisferi, ai tropici, sulle montagne, nei paesi
nordici, ma anche in Nuova Zelanda, con diversi e numerosi generi. Hanno fusto
elevato, a ramificazione monopodiale, con accrescimento apicale, superante in
vigore e altezza quello laterale. Anche nel pino ad ombrello, abbiamo sempre un
punto di meristema, più alto di tutti; il tipo monopodiale per eccellenza,
resta però l’abete.
Abbiamo macroblasti e brachiblasti, i primi portanti
i secondi e i secondi, portanti le foglie. In Cipresso e Cryptomeria, invece, le
foglie sono portate direttamente sul fusto; nel Cedro sono ben evidenti i
brachiblasti. Il protoxilema è endarco, cosa che si avvicina alle Dicotiledoni
nella struttura primaria a fasci collaterali. Come struttura secondaria, abbiamo
profonde trasformazioni, con l’insorgere di un cambio interfasciale, che dà
un anello continuo di cambio, che proliferando, dà anelli concentrici di floema
e xilema. Questa struttura è presente in tutte le Conifere, dato che sono tutte
perenni, sia le altissime Sequoia, sia le basse, come lo Juniperus montana, sia
striscianti sul terreno, come il Podocarpus australe.
Esaminiamo una struttura secondaria in sezione
trasversale, longitudinale tangenziale e longitudinale radiale.
Nel primo caso è possibile vedere il contatto fra il
legno di primavera e l’autunnale, per la differenza delle cellule. Il legno di
primavera e abbondante, a cellule grandi, con tracheidi a lume molto grande.
Quando l’umidità diminuisce e si va verso la stagione secca, diminuisce il
lume delle tracheidi. Si nota una perfetta concordanza fra struttura del legno e
clima.
Nella sezione, si possono notare i raggi midollari,
parenchimatici, di diversa lunghezza e visibilità, a seconda del loro ordine e
del momento di formazione. Si nota pure il canale resinifero, con una parte
centrale lacunare, circondata dalle cellule secernenti e più esternamente, da
cellule difensive, per impedire lo schiacciamento.
Nella seconda sezione sono evidenti le abbondanti
tracheidi, il canale resinifero, molte strutture che appaiono lenticolari e che
non sono altro che i raggi midollari tagliati. Sono fatti di cellule
parenchimatiche e talvolta racchiudono al centro un canale resinifero; le
cellule parenchimatiche hanno, caratteristicamente un allungamento in senso
radiale. Presenti, sono le punteggiature, che sono le aperture che permettono il
collegamento fra una tracheide e quella vicina. Sono strutture già descritte
nella parte generale ed essenzialmente formate da una lamella che separa le
tracheidi tra loro. Intorno alla punteggiatura, vi è un ispessimento di
cellulosa, detto cercine, che conferisce alla cellula un aspetto a coda di
rondine, con in mezzo la lamella mediana, che ha la parte centrale ispessita,
questa è detta toro. Diverse sono le ipotesi sull’utilizzazione di
questo complesso. In condizioni normali, quest’insieme permette normalmente il
passaggio di sostanze da una tracheide all’altra, ma qualora si necessiti di
una maggiore spinta d’ascesa, il toro, cioè la parte centrale ispessita della
lamella mediana fa da valvola, andando ad ottundere la perforatura di una delle
due tracheidi, cosicché l’acqua passa per una sola delle due, e il liquido
riceve maggiore spinta.
Nella terza sezione si possono vedere le tracheidi
con le punteggiature che hanno ora un aspetto tondeggiante dato il taglio, il
canale resinifero, il raggio midollare di cellule allungate in senso radiale e
di cellule parenchimatiche, dirigentisi alla periferia. Questa sezione è di
difficile preparazione perché è assolutamente necessario che passi per il
centro esatto.
Posseggono midollo piccolo, in accordo con la loro
altezza e la vita in ambienti non molto ospitali.
Le foglie sono inserite o tutte sui rami stessi o
tutte sul brachiblasto; nel primo caso con disposizione decussata o
verticillata, nel secondo caso a spirale. Sono aghiformi, lineari, talvolta
corte come nel Cedro, squamiformi nel Cipresso, più espanse nel Podocarpus.
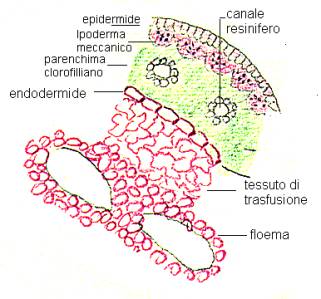
Eseguiamo l’anatomia della foglia più tipica:
l’ago di pino.
Dall’esterno troviamo un’epidermide,
riconoscibile dalla mancanza di cloroplasti, ricoperta da uno strato spesso ed
abbondante di cutina. Sotto di essa, troviamo uno strato ipodermico o ipoderma
meccanico, fatto da cellule di sostegno, fibriformi, allungate in senso
parallelo alla nervatura. Sono cellule a parete ispessita, perciò
sclerentimatiche. All’interno, abbiamo ancora, la vera e propria parte
parenchimatica clorofilliana, ricca di cellule con cloroplasti, la cui parete
s’introflette per brevi tratti; le pareti delle cellule vicine, sono
introflesse in posizione corrispondente e opposta, si realizza in questo modo,
una maggiore aerazione, all’interno del parenchima clorofilliano, si vedono le
tracce delle nervature, individuate da uno strato esterno detto endodermide. È
fatto da una sola fila di cellule, a disposizione circolare. Nel suo interno,
troviamo il tessuto di trasfusione, esclusivo del gruppo, fatto da parenchima di
riserva e aquixilema, ricchi di sostanze di riserva contenute in cellule
albuminifere, che devono il nome all’aspetto delle sostanze presenti,
dall’aspetto simile all’albume d’uovo. All’interno del tessuto di
trasfusione, troviamo i fasci cribro-legnosi, accoppiati con floema e xilema.
Spesso, intercalati perpendicolarmente a questo parenchima, troviamo canali
resiniferi di tipo schizogeno, cioè fatti da cellule sempre secernenti.
Gli strobili sono unisessuali, cioè a sessi separati, ma portati sulla stessa pianta. I maschili sono fatti da brattee a funzione protettiva e da squame fertili portanti le sacche polliniche (sono presenti in numero di 2-20). Sono portate nella parte dorsale della squama, cioè nella pagina inferiore, per facilitarne la dispersione. All’interno delle sacche si producono le microspore, che poi si trasformano in polline. Quando il polline si distacca, non è più considerato microspora, ma già gametofito maschile, perché in esso, abbiamo generalmente, più di un nucleo. Il trasporto del polline è facilitato dalle due sacche aerifere vuote, laterali, nate per estroflessione della parete e che servono per aumentare la superficie di galleggiamento. Portato dal vento sul micropilo della parte femminile, penetra nella camera pollinica. Esso possiede ora un nucleo e anche una o due cellule protallari piccole ed un’altra di grandi dimensioni che subito si divide in due, dando la cellula vegetativa a funzione trofica e la cellule anteridiale, deputata alla riproduzione.
La cellula vegetativa produce il tubetto pollinico che s’accresce enormemente, penetra attraverso la nucella, si ramifica abbondantemente per assorbire nutrimento e viene a contatto con l’oosfera. È ormai un tubetto pollinico vettore.
La cellula anteridiale dà una cellule parietale e una cellula madre degli spermi, che darà i nuclei spermatici, in numero di due, entrambi utilizzati, per cui si parla già di doppia fecondazione.
Gli strobili femminili sono molto piccoli da giovani,
mentre da adulti assumono grandi dimensioni e sono ciò che noi chiamiamo pigne.
Si accrescono in tre anni, poi cadono. Sono fatti da brattee sterili e squame
fertili, il tutto inserito sull’asse dello strobilo, a spirale. La brattea sta
sotto, la squama è attaccata sopra ad essa. Talvolta possono essere
completamente saldate fra loro, e talvolta completamente fuse. Le brattee al
momento opportuno si aprono, allorché giunge il granello pollinico, poi si
richiudono e restano così fino al momento della disseminazione.
La squama porta gli ovuli, ortotropi, ma nel corso
dell’evoluzione si può avere un ribaltamento verso il basso, non dovuto mai
al ribaltamento del supporto dell’ovulo, per cui questo non è anatropo, ma
sempre ortotropo; la posizione verso il basso è dovuta al ribaltarsi della
squama.
Nell’ovulo abbiamo dall’esterno: tegumento,
micropilo, camera pollinica, nucella, cellula madre delle spore, che dà origine
a quattro spore, di cui una sola matura e darà il gametofito femminile o
protallo o endosperma primario di cellule apolidi a funzione esclusivamente
trofica. Troviamo in esso due oosfere, sormontate dai soliti residui
archegoniali di canale e ventre; le oosfere possono essere anche più numerose.
Con la fecondazione, n’è interessata una sola, e se più, l’embrione è
sempre uno, le altre cellule degenerano. La fecondazione è semplice, è
utilizzato un solo nucleo spermatico, ma si è visto, in rari casi, che la
fecondazione può essere doppia, cioè uno dei due nuclei feconda l’oosfera e
l’altro va ad unirsi alle cellule residue dell’archegonio; abbiamo così, un
insieme diploide che prolifica e dà cellule a funzione trofica. È interessante
perché preludio alla comune e normale fecondazione delle Angiosperme. Dalla
fecondazione si ha poi l’embrione.
Tra l’impollinazione e la fecondazione, possono
passare anche varie settimane.
Gli strobili cosa rappresentano filogeneticamente ?
Si è affermato che corrispondano perfettamente a quelli delle Licopsida, oggi
però si preferisce interpretarlo come qualcosa che corrisponde ad una sorta
d’infiorescenza. Esaminiamo la parte femminile.
Si è visto che nei fossili si aveva un asse centrale
portante alla base una brattea, ma nell’ascella si ha un ramoscello che porta
la squama, nella parte inferiore di essa c’è l’ovulo.
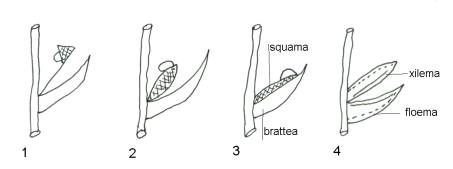
Ad un certo punto (2), l’asse del ramoscello è
scomparso e la squama è direttamente inserita nell’ascella. In (3) la squama
si è completamente saldata alla brattea; si è così raggiunta la condizione
moderna con un processo d’annullamento seguito da saldatura. L’ovulo che
vediamo sopra la squama, e perciò in apparente posizione contraria rispetto la
condizione del maschio, è frutto di uno slittamento e in definitiva ovuli e
sacche polliniche hanno la stessa posizione, inferiormente alla squama. È uno
slittamento filetico, per cui pur mantenendosi sempre la stessa topografia, si
è raggiunto un completo sistema protettore.
Anche per l’anatomia, le cose corrispondono;
infatti, nella squama, la parte floematica (tratteggiata), è situata
superiormente, la xilematica inferiormente, a contatto con lo xilema della
brattea, che ha il floema nella sua parte inferiore, come nella foglia.
I semi maturano lentamente, in genere ci vuole più
di un anno, il che permette la comparsa di un nuovo ramo; anzi talvolta gli anni
sono tre e troviamo tre pigne, d’età diverse, inserite sullo stesso ramo, la
più vecchia in basso, la più giovane in cima. Ciò denota una notevole
lentezza nel reagire ai processi indotti dalla fecondazione. Spesso il seme è
alato; le ali sarebbero trasformazioni della parte periferica del seme stesso o
deriverebbero dalla squama fertile. Ciò che è interessante è che nel seme
abbiamo tegumenti seminali, endosperma primario ed embrione; i tegumenti
appartengono alla generazione sporofitica primaria, l’endosperma è di natura
gametofitica, l’embrione è lo sporofito secondario. È importante perché
nelle Angiosperme avremo tegumenti ed embrione, ma non più la parte
gametofitica, perché qui la parte trofica insorge a seguito della doppia
fecondazione. L’embrione può avere uno o più cotiledoni, rappresentati dal
ciuffettino di foglie in cima ai pinoli appena nati.
I coni o strobili fruttiferi femminili sono
caratteristici da gruppo a gruppo (es: pino, cedro, sequoia, cryptomeria,
cipresso); a maturità si aprono e lasciano cadere i semi. Solo nel ginepro le
“palline” restano chiuse; in esse le squame e le brattee sono carnose e
diconsi galbuli.
Comprendono vari ordini:
Lebachiales: fossili del Carbonifero, rari.
Voltziales: fossili, ben rappresentati fino al Mesozoico, molto
comuni.
Pinales: possiamo ricordare diversi esemplari attuali che
raduniamo nelle famiglie delle
Taxodiaceae: fra queste si ricordano le Sequoia. La Sequoia
dell’orto botanico dell’università di Genova è stata piantata nel 1835.
Sono fra le piante più longeve, sembra oltre 4000 anni. Anche il Taxodium è
fra le Conifere messicane più antiche. La Metasequoia si ricorda come esempio
di fossile vivente, infatti, era un fossile di comune ritrovamento fino
all’Eocene. Si riteneva estinta, quando fu scoperta in una valle cinese con
circa 100 esemplari, in tutto simili ai fossili. I semi furono piantati in
diversi orti botanici e parchi. Come specie e dystica, se ci si riferisce ai
fossili, cryptostroboides se attuale.
Pineaceae: cui appartengono i generi Abies, Larix, Pinus,
Cedrus. Quest’ultimo in genere coltivato, perché spontaneo solo in Himalaya.
L’Abies è spontaneo da noi come Abete bianco, appenninico; in Sicilia c’è
l’Abies nebrodensis. Da ricordare che tutti gli abeti mediterranei, pur
essendo molto diversi morfologicamente, sono interfecondi, per cui siamo di
fronte a tanti singameoni. L’abete rosso o Picea abies o Picea excelsa, dai
rami penduli, è comune sulle Alpi; restano rari gli esemplari sull’Appennino
tosco-emiliano. Del genere Pinus, numerose sono le specie; il Pinus pinea o pino
da pinoli o pino domestico o pino italico forma le pinete di Viareggio, di
Ravenna, di S.Rossore, di Migliarino; il Pinus cembra con cinque aghi per
brachiblasto, il Larice con foglie decidue e presente sulle Alpi anche ad alte
altitudini.
Cupressaceae: ricordiamo lo Juniperus phoenicea o cedro licio o
ginepro fenicio che presenta il fenomeno dell’eterofillia, con foglie
aciculari o squamiformi, in relazione allo sviluppo; il Cupressus sempervirens,
il Cupressus macrocarpa (California), il Cupressus lusitanica (Messico), il
Cupressus arizonica (Arizona).
Cephalotaxaceae: rappresentante è il Podocarpus che ha
strobili ridotti a due soli ovuli.
Sono ben distinte dalle altre conifere. Il Taxus baccata è l’unico rappresentante in Italia. È un albero, talora arbusto, molto longevo che risale al Triassico superiore. La più grande differenza con le Conifere è nella mancanza di canali resiniferi.
Le foglie sono, distiche, lineari, falcate con inserimento a spirale o decussato, senza differenza fra macro e brachiblasti; per torsione però, le foglie appaiono disposte in un piano, con una specie di pettine dai due lati del rametto.
Gli strobili sono sempre all’ascella delle foglie;
il femminile, ha un solo ovulo apicale. I maschili hanno brattee basali
decussate e lungo l’asse s’inseriscono i microsporofilli che possono essere
di due tipi: dorsoventrali isosporangiati, se fatti da squame che sotto
portano i macrosporangi; peltati perisporangiati se sono a mo’ di scudo
con ai margini le sacche polliniche. La differenza non ha valore sistematico. Il
polline non ha sacche; la fecondazione avviene per mezzo di nuclei spermatici,
diversi per dimensione, uno grosso utilizzato ed uno più piccolo; è esempio
verso una tendenza ad una specializzazione della fecondazione, diversamente che
nelle Coniferopsida.
La parte femminile è molto ridotta. Si pensa che in
origine si avessero dei veri strobili, anche se ora il tutto è ridotto al solo
ovulo, isolato, ortotropo, cioè eretto, cosa che ci ricorda il Ginkgo, solo che
qui avevamo due ovuli. Ora abbiamo un solo ovulo con supporto e quindi, è un
modello che si è ancora più ridotto.
Una volta fecondato, l’ovulo si trasforma in seme;
via via che avviene la maturazione, attorno all’ovulo compare un anello
carnoso rossastro, prodotto dall’asse di sostegno dell’ovulo e che lo
avvolge da tre lati: è chiamato arillo. La disseminazione è ornitocora;
infatti, l’arillo, ricco di sostanze nutritizie e di colore rosso, è
appettito dagli uccelli che, cibandosene ne favoriscono la disseminazione. Da
notare che la pianta è velenosa, mentre l’arillo è edule. La veneficità è
la ragione per cui sia poco diffusa, essendo stata abbattuta nei luoghi di
pascolo.
Le Taxopsida hanno affinità con le Coniferophytina,
pur essendo a sé stanti per il biochimismo (velenosità), per la mancanza di
sacche polliniche; si considerano nate per riduzione dalle Coniferopsida, più
antiche. Già nelle Conifere si aveva riduzione degli strobili (Ginkgo)
femminili, qui è molto più spinta.
Le Coniferophytina, sono in definitiva un gruppo ben
“affiatato”, ma con quattro classi ben distinte, dapprima con fecondazione
per mezzo di spermatozoi, poi con fecondazione per nuclei spermatici. Da tenere
presente, ad ogni modo, che non sono classi sullo stesso piano e nemmeno le une
discendenti dalle altre, ma sono derivate da dicotomie successive di una linea
originaria comune. Il progenitore è ignoto e anche quali siano gli effettivi
rapporti filogenetici fra loro. Un antenato potrebbe ricercarsi fra le Licopsida,
cosa che non potrebbe essere contraddetta, ma neppure confermata; in ogni caso
sono da escludersi dalla linea filetica delle Conifere, le Cycadophytina e le
Pteridospermophytina, per cui la scelta, ci resta o fra le Licopsida, o in un
essere ignoto.
Hanno apparati sporofillici con foglie perianziali,
cioè hanno i fiori. È un fiore primitivo, con un abbozzo di perianzio.
Il legno secondario è eteroxilo, qui compaiono per
la prima volta le trachee (in alcune felci, ci sono casi d’eteroxilia).
I macrosporofilli sono molto trasformati, fatti da
entità isolate o costituenti delle infiorescenze; anche i microsporofilli sono
diversi da quelli finora visti.
La fecondazione avviene per nuclei spermatici.
Sono arbusti, suffrutici, liane, sarmenti, fusti
volubili ecc.; mancano dello stadio arboreo. Il tronco più alto è nella
Welwitschia, dove arriva a 1- 1,5 m. le foglie sono opposte verticillate,
talvolta enormi ad accrescimento continuo.
I fiori sono unisessuati, cioè ogni fiore ha un suo
sesso. Questo è un fatto che porta il problema, se lo stato dioico sia più
primitivo o no dello stadio monoico.
I fiori maschili costituiscono infiorescenze,
ciascuno è costituito da un perianzio di due pezzi, con le sacche polliniche
saldate insieme fra loro a costituire dei sinangi. Troviamo anche dei sinandri,
perché più sinangi possono saldarsi fra loro in una nuova entità, il
sinandrio.
Il fiore femminile ha una parte perianziale propria,
con uno o due ovuli, ciascun portatore di due tegumenti, di cui il più interno
è provvisto di una colonna micropilare che sporge fuori del tegumento più
esterno. È una facilitazione per la cattura del granello pollinico. Siamo però
in presenza, ancora di un ovulo, non di un ovario.
Dall’ovulo deriva il seme, poi l’embrione con due
cotiledoni; questo ci dimostra che la riduzione numerica dei cotiledoni è un
fatto che procede con l’evoluzione, per cui le monocotiledoni sono le piante
più evolute.
Lo sviluppo delle foglie ha messo in chiaro una
differenza sostanziale del gruppo. Così nelle Ephedrales abbiamo un solo
meristema apicale definito che dà foglioline minute (solo apice precursore);
nella Welwitschia invece, siamo in presenza di un meristema basale indefinito,
responsabile delle foglie larghe 15-20 cm. che si dipartono a destra e a
sinistra del tronco e crescono come due lunghissimi nastri.
Volendole inserire in una linea filogenetica, le cose
si complicano. Hanno caratteristiche primitive nettissime, ma non abbiamo
fossili. Sono vicine sia alle Conipherophytina sia alle Angiosperme, ma la cosa
più probabile è che siamo di fronte ad una linea conservatrice laterale, come
per lo Psilotum. La teoria della linea conservatrice è appoggiata dall’aversi
tre generi distinti, sparsi in diverse regioni del globo e così distinti che
c’è chi le considera tre classi o addirittura due, con Gnetum ed Ephedra
insieme e separata le Welwitschia. Noi consideriamo una sola classe e
distinguiamo in essa tre ordini: Welwitschiales con la Welwitschia, dalle enormi
foglie nastriformi e per il meristema basale indefinito; le Ephedrales con l’Ephedra,
dalle foglie minutissime, quasi invisibili, ridotte al solo apice precursore; le
Gnetales con lo Gnetum.