
|
Riassunto: La questione dell'asimmetria molecolare degli amminoacidi costituisce un problema la cui soluzione comprende due aspetti. Infatti, chiarire se l'origine dell'asimmetria in questione sia una condizione per la comparsa della vita sulla terra o una sua conseguenza, equivale a considerare la vita un evento molto raro, che puň anche essere irripetibile, oppure la inevitabile evoluzione di alcune strutture molecolari in un ambiente di reazione molto particolare.
Nel primo caso, sono stati proposti alcuni processi fisici (luce polarizzata, effetti del campo magnetico terrestre, cristallizzazioni asimmetriche) per spiegare l'origine dell'asimmetria in questione, ma le prove sperimentali non sono ritenute soddisfacenti in quanto relative a condizioni estremamente severe e particolari, caratterizzate da riproducibilitŕ praticamente nulla. Piů stimolanti sembrano le ipotesi che vorrebbero spiegare la possibilitŕ di sintesi asimmetriche mediante reazioni regolate da cinetiche del non equilibrio; tuttavia, queste ipotesi non sono del tutto soddisfacenti in quanto le reazioni ipotizzate, qualora si fossero verificate, sarebbero verosimilmente avvenute contemporaneamente ed in molti punti del brodo primordiale, generando in media ancora una soluzione racemica. Le ipotesi riassunte, comunque, accordano un'importanza forse troppo grande all'aspetto strutturale della vita, a detrimento del suo aspetto funzionale. Da un altro punto di vista, con l'uso di modelli matematici, comunemente utilizzati per analizzare e spiegare il comportamento di semplici ecosistemi, č possibile tracciare uno schema d'insieme che puň fornire una spiegazione dell'asimmetria biologica nel quadro della selezione naturale. |
La prima ipotesi biologica (1957) č dovuta a George Wald, il quale, basandosi sulla considerazione che gli acidi nucleici e le proteine di ogni singolo organismo comunque complesso debbano essere stereochimicamente compatibili, propose che le prime unitŕ prebiologiche originatesi in seno al brodo primordiale, potessero costituire due distinte popolazioni enantiomorfe. Successivamente, nel corso dell'evoluzione, una delle due popolazioni avrebbe acquisito casualmente sull'altra un qualche vantaggio evolutivo e conseguentemente una certa prevalenza nella lotta per l'esistenza. Questa ipotesi, tenendo conto che i vari organismi si nutrivano uno dell'altro, scambiandosi materiali attraverso complesse reti alimentari, permetterebbe di darsi ragione di come, sotto la pressione del processo evolutivo, si possa essere selezionata ubiquitariamente sulla Terra la vita a base di soli L-amminoacidi.
In realtŕ, non č facile immaginare - né Wald lo suggerisce - come una delle due popolazioni enantiomorfe possa aver acquisito globalmente un certo vantaggio sull'altra. Infatti, le unitŕ prebiologice che costituivano dette popolazioni, erano probabilmente caratterizzate da notevole mutabilitŕ, sicché le numerose variazioni che si presentavano fra i vari individui ad essi appartenenti, verosimilmente si compensavano, di modo che, sebbene alcune specie si siano potute estinguere, č pensabile che globalmente le due popolazioni percorsero la loro evoluzione in modo non dissimile, comunque tale da rendere discutibile la possibilitŕ che abbiano potuto promuovere significativi processi di selezione stereochimica.
La possibilitŕ che le due popolazioni enantiomorfe abbiano avuto un'evoluzione parallela, ha suggerito la possibilitŕ che il processo di selezione dei L-amminoacidi sia avvenuto quando comparvero i primi organismi interessati a scambi di materiale genetico oppure a riproduzione sessuata. Secondo questa ipotesi, si sarebbero potute sviluppare, fino a questo stadio, due popolazioni del tutto simili il cui unico svantaggio dal punto di vista evolutivo sarebbe stata la sterilitŕ degli accoppiamenti misti: questa sarebbe stata la causa dell'estinzione di alcune popolazioni.
Per concretizzare la questione, indichiamo con x ed y il numero di individui di un sesso (ad es. femmine) appartenenti rispettivamente alla prima ed alla seconda popolazione. Detti individui, col tempo, sono soggetti a due opposte sorti: tendono ad aumentare per conseguenza degli accoppiamenti compatibili ed a diminuire a causa della mortalitŕ naturale. Il modello matematico proposto per descrivere l'evoluzione di dette popolazioni enantiomorfe č il seguente sistema di equazioni differenziali:
dove k e c sono due coefficienti di proporzionalitŕ che misurano rispettivamente i tassi di natalitŕ e mortalitŕ. In particolare, si č qui supposto che il numero di accoppiamenti per unitŕ di tempo cui partecipavano le femmine della prima popolazione sia proporzionale al loro numero x, ma la sola frazione fertile di tali accoppiamenti č x/(x+y), corrispondente al rapporto tra gli accoppiamenti compatibili (con maschi della stessa simmetria) e quelli possibili (anche con maschi della simmetria opposta). La mortalitŕ per unitŕ di tempo tra le femmine della prima popolazione č assunta proporzionale al numero delle femmine stesse.
In modo analogo si formula la seconda equazione differenziale del sistema (1) i cui integrali generali - supposti costanti i coefficienti k e c - sono:
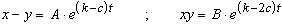
con A e B costanti arbitrarie di integrazione.
Ora, escluso il caso in cui A = 0, cioč corrispondente ad una uguale consistenza numerica delle due popolazioni, in quanto questa situazione non č compatibile con le inevitabili fluttuazioni statistiche nel numero di individui della stessa popolazione, possiamo senz'altro discutere le soluzioni.
a) k < c - le due funzioni esponenziali tendono a zero per 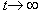 . Questo significa che entrambe le popolazioni si estinguono in quanto il tasso di mortalitŕ č maggiore del tasso di natalitŕ. In particolare, poiché la differenza numerica tra le due popolazioni tende ad annullarsi, la popolazione piů numerosa ha un tasso di mortalitŕ piů elevato.
. Questo significa che entrambe le popolazioni si estinguono in quanto il tasso di mortalitŕ č maggiore del tasso di natalitŕ. In particolare, poiché la differenza numerica tra le due popolazioni tende ad annullarsi, la popolazione piů numerosa ha un tasso di mortalitŕ piů elevato.
b) k >> 2c - le due funzioni esponenziali divergono per . Questo significa che entrambe le popolazioni crescono illimitatamente, ma la piů numerosa con un ritmo maggiore in quanto anche la loro differenza diverge.
c) c<k<2c delle due funzioni esponenziali, per , la prima diverge e la seconda converge. Questo significa che la popolazione piů numerosa tende a crescere piů rapidamente, mentre l'altra si estingue: nella seconda equazione č sufficiente che una delle due variabili (x, y) si annulli, quella corrispondente alla specie meno numerosa.
Possiamo escludere il caso (b) in quanto le equazioni (1) costituiscono un modello che conserva la sua validitŕ solo per un intervallo di tempo limitato. Infatti, alla fine una popolazione in continua crescita esaurirŕ le proprie risorse vitali; questa limitazione si puň considerare aggiungendo al sistema (1) il cosiddetto smorzatore: un termine sottrattivo rappresentante le limitazioni nutrizionali. D'altra parte, per non appesantire il modello, si puň ragionare qualitativamente, osservando che i coefficienti k e c non sono realmente costanti, bensě funzioni del tempo e della densitŕ delle popolazioni. Sicché, posto k>2c, allorché la densitŕ di popolazione raggiungerŕ un valore critico legato alle disponibilitŕ nutrizionali, k(natalitŕ) inizierŕ a diminuire e c (mortalitŕ) a crescere fino a raggiungere la condizione c<k<2c.
Cosě, sembrerebbe dimostrato che la popolazione piů numerosa tende ad un valore limite per cui k = c, mentre l'altra popolazione č condannata all'estinzione. Pertanto, riallacciandoci a quanto ipotizzato da Wald, si potrebbe anche spiegare come lo stesso tipo di simmetria si sia imposto su tutta la biosfera: le complesse reti alimentari fecero sě che anche gli organismi a riproduzione asessuata, dovendosi nutrire dei resti di organismi piů complessi, vennero selezionati giacché erano favoriti quelli che poterono inserirsi nel ciclo stabile della popolazione enantiomorfa superstite.
In realtŕ, le equazioni proposte non costituiscono il supporto di una spiegazione soddisfacente, in quanto sia a livello di riproduzione sessuata tra forme piů complesse (alga Chlamydomonas, alcuni batteri), sia a livello di scambi di materiale genetico tra forme prebiotiche, ammettono implicitamente che anche a seguito di un "accoppiamento" sfavorevole venga saltata una "stagione di riproduzione" ossia un ciclo riproduttivo e non prevedono la possibilitŕ che il materiale genetico incompatibile venga eliminato tout court.
| Si può offrire una sorta di conferma sperimentale a questa critica. Sono state sperimentate tecniche d'impiego di feromoni per evitare la diffusione di parassiti senza ricorrere a sostanze tossiche. I feromoni, vaporizzati nell'ambiente, inducono gli insetti maschi ad accoppiarsi anche con femmine immature che quindi non genereranno. I risultati si sono dimostrati interessanti solo se la densitŕ di popolazione dei parassiti è scarsa e le specie sono residenti. |
Inoltre, secondo tale processo di selezione sessuale, la popolazione piů numerosa tende ad eliminare quella meno numerosa poiché gli accoppiamenti misti lasciano disponibile un maggior numero di elementi appartenenti alla prima popolazione, e questi possono riprodursi. Ciň perň č ragionevole per due popolazioni costituite ciascuna da un numero di maschi uguale a quello delle femmine e aventi una eguale disponibilitŕ di alimenti asimmetrici delle due forme. Pertanto, premesso che una elaborazione del sistema (1) tale da eliminare le critiche esposte porterebbe ad una sorta di simulazione matematica di difficile se non impossibile soluzione, ed in ogni caso le soluzioni sarebbero di non facile interpretazione, rimane comunque il fatto che la teoria cosě proposta costituisce solo una possibilitŕ.
D'altra parte, poiché sulle formule non č scritto il loro campo di applicazione ma siamo noi che facciamo qualcosa con esse, possiamo interpretare il sistema (1) come la descrizione temporale di un particolare sistema predatore-preda in cui due popolazioni enantiomorfe si muovono in modo arbitrario cibandosi l'una dell'altra. Quando due individui ad esse appartenenti si incontrano, esiste una probabilitŕ costante che uno dei due uccida l'altro impiegando un tempo trascurabile. In particolare, per l'occasionale predatore, la preda stereochimicamente incompatibile sarebbe indistinguibile dall'altra, sicché cibandosene trarrebbe un nutrimento limitato alle sole molecole simmetriche. Questa interpretazione - che a differenza della precedente - prevede che in ogni scontro un individuo sia eliminato con possibile vantaggio per l'occasionale predatore - richiede l'ipotesi che il cibo costituito da molecole simmetriche sia illimitato in confronto a quello asimmetrico ricavabile essenzialmente dalla predazione; non č comunque necessario che il numero dei maschi e delle femmine appartenenti a ciascuna popolazione sia uguale.
Da quanto discusso, viene sottolineato come al proposto processo di estinzione di una delle due popolazioni enantiomorfe per incompatibilitŕ di materiale genetico, si possano sovrapporre le esigenze nutrizionali che condurrebbero inevitabilmente ad una sorta di competizione. Competizione passiva, č vero, ma tuttavia competizione in quanto certi sistemi sopravvissero ed altri scomparvero per sempre.
Si presenta quindi la necessitŕ di discutere il comportamento di un sistema competitivo. Come premessa, č bene sottolineare che perché si verifichi competizione, non č necessario che scarseggino tutti gli elementi nutritivi: č sufficiente che spariscano o si rendano scarsamente disponibili alcuni componenti fondamentali. Quando ciň si verifica, come fatto del tutto accidentale, viene favorita una delle due specie.
Per questa discussione, ci baseremo sulle equazioni di Lotka-Volterra, ma anziché proporre possibili vantaggi evolutivi responsabili della selezione dei L-amminoacidi, č piů conveniente invertire i termini della questione esaminando quali condizioni debbano essere soddisfatte, in linea di principio, perché due popolazioni enantiomorfe in competizione possano coesistere stabilmente.
Faremo l'ipotesi che dette popolazioni ad un certo punto della loro evoluzione si diversifichino, senza per questo affermare che una delle due abbia un vantaggio sull'altra. Siano:

le equazioni che descrivono l'evoluzione temporale di due specie in competizione. con a, b, c, e, f, g, costanti positive il cui significato č il seguente:
a, e : costanti che misurano l'indice di natalitŕ;
b, f : costanti che misurano le limitazioni nutrizionali (quando il numero di individui x o y č basso, i termini sottrattivi bx2 ; fy2 sono trascurabili e la crescita č esponenziale);
c, g : costanti che misurano gli effetti inibitori di ciascuna specie sul proprio competitore.
Facciamo l'ipotesi che il fattore critico limitante la possibile coesistenza delle due popolazioni sia essenzailmente legato alle limitazioni nutrizionali; ciň equivale ad imporre:
risolvendo il sistema, si presentano due casi interessanti:
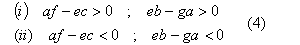
Con le condizioni (i), si puň dimostrare che il sistema č stabile, mentre con le (ii) č instabile: quale delle due popolazioni prevarrŕ dipende dalle condizioni iniziali. In effetti, il secondo caso (ii) corrisponde alla formulazione in termini analitici della teoria proposta da Wald e valgono evidentemente le stesse critiche; rimane dunque da discutere il primo caso (i).
Per interpretare biologicamente la condizione (3) relativa alla coesistenza delle due popolazioni enantiomorfe, supponiamo, senza diminuzione di generalitŕ, che le velocitŕ intrinseche di accrescimento a, e delle due popolazioni siano uguali, dunque la condizione espressa dalle (4) risulta : b>g e f>g. Queste disuguaglianze stabiliscono che un aumento di densitŕ di una delle due popolazioni comporterŕ una limitazione del proprio accrescimento maggiore di quella indotta nella popolazione competitrice. Questa eventualitŕ puň verificarsi se due popolazioni sono limitate parzialmente da differenti risorse vitali: puň cosě essere arrestata la crescita della popolazione caratterizzata da maggior tasso di accrescimento prima che raggiunga una densitŕ sufficiente a minacciare l'estinzione dell'altra. Nel caso in esame, la coesistenza di due popolazioni enantiomorfe puň essere favorita proprio dalla impossibilitŕ di una comune utilizzazione di molecole asimmetriche.
Ovviamente, si pone un altro problema: per quanto tempo potrŕ mantenersi questa situazione di stallo? Non bisogna infatti dimenticare che un modello deterministico come quello proposto non puň riflettere la realtŕ ecologica in quanto presuppone come infinite le dimensioni della popolazione e trascura i fattori casuali che si verificano col tempo nell'ambiente circostante.
Cosě, ad esempio, se un modello deterministico prevede che la densitŕ di una o piů specie in un ecosistema scenda periodicamente a valori molto bassi, il corrispondente modello stocastico, invece darŕ una probabilitŕ non trascurabile di estinzione. D'altra parte, se un modello deterministico indica un equilibrio stabile, il corrispondente modello stocastico prevederŕ una sopravvivenza a lungo termine.
Comunque, pur con queste limitazioni, l'applicazione di un qualsiasi ragionevole modello matematico riesce utile in quanto non č necessario che fornisca una descrizione precisa e rigorosa della corrispondente realtŕ biologica. Infatti, ciň che vogliamo da un modello matematico č conoscere quale tipo di variazione nel comportamento del modello sia indotta da una particolare variazione dei parametri, e poter dedurre se una corrispondente variazione si possa essere verificata nel comportamento del sistema in questione. Cosě, nel nostro caso, con il modello proposto ci siamo posti nella condizione diversa da quella offerta dalla teoria di Wald: č individuato un possibile fattore stabilizzante.
Ciň premesso, possiamo formulare l'ipotesi che una volta raggiunta una situazione di stallo, si sia verificata una selezione fra le varie specie appartenenti a ciascuna popolazione, fino a ridurre la competizione essenzialmente a due specie enantiomorfe, delle quali una si estinse. Questo punto di vista puň essere avvalorato dalle proprietŕ di autorganizzazione dei proteinoidi di Fox e dai meccanismi di selezione delle microgocce di Oparin: i composti chimici che in uno stesso sistema sono coinvolti in reazioni parallele, sono trasformati nella direzione ove agiscono piů rapidamente. Cosě, in un organismo vivente o prebiotico, quelle reazioni che avvengono lentamente, potranno difficilmente inserirsi in un ciclo metabolico: questa selezione si ripercuote evidentemente in tutto il sistema. Pertanto, in un ambiente chiuso dove miriadi di organismi primitivi coesistono e sono in competizione passiva, il flusso piů significativo di materia ed energia attraverserŕ il sistema piů efficiente, ossia quello che possiede la maggior velocitŕ di trasformazione chimica interna. Se il loro dinamismo č compatibile con l'autoconservazione, questi sistemi sottrarranno una significativa quantitŕ di sostanze organiche ai sistemi caratterizzati da metabolismo piů lento che verranno via via eliminati. Solo gli organismi piů dinamici, caratterizzati da catalizzatori piů efficaci, sono stati conservati dalla selezione naturale.
Sulla base di quanto discusso, non č chiaro come avvenne questa selezione; possiamo solo accennare alcune possibilitŕ.
Se ad esempio formuliamo il sistema (2) ponendo a = e; c = g; bf -c2>0, ossia consideriamo due specie enantiomorfe con velocitŕ intrinseche di accrescimento uguali ma tali da limitare l'accrescimento in misura maggiore da quanto imposto da una differente consistenza di sostanze nutritive asimmetriche (c > b > f), allora si vede immediatamente che una delle due specie si estingue.
Questo modello (o meglio dette condizioni), puň servire da traccia per spiegare l'utilizzazione da parte dei sistemi viventi di soli a amminoacidi. Infatti, nelle sintesi realizzate in laboratorio, la proporzione degli alfa amminoacidi č superiore a quella dei beta amminoacidi; ciň suggerisce che allorché si formarono i primi polipeptidi, gli a amminoacidi fossero piů abbondanti dei b amminoacidi, sicché si verificň una prima selezione fra i sistemi prebiotici che si trovavano in competizione passiva: sopravvissero quelli capaci di utilizzare il gruppo di amminoacidi piů abbondante. Allo stesso modo, una fluttuazione nella consistenza numerica dei L e D amminoacidi potrebbe aver operato la selezione in causa. In questo caso, si noti che non č necessario che tutta la classe dei L amminoacidi sia stata piů numerosa dell'altra: č sufficiente che alcuni D amminoacidi siano stati meno diffusi e numerosi. Analogo destino subiscono due specie enantiomorfe le cui velocitŕ intrinseche di accrescimento siano uguali a = e ma i cui reciproci effetti inibitori siano differenti e limitino l'accrescimento in misura maggiore di quanto imposto da una eguale consistenza di sostanze nutritive asimmetriche (c > b = f).
L'osservazione che alcuni antibiotici naturali (gramicidina S, penicillina) devono forse in parte la loro attivitŕ all'alto contenuto della configurazione D degli amminoacidi, suggeri-ce anche una formulazione alternativa del sistema (2) in modo da considerare la possibilitŕ che ogni specie enantiomorfa produca una sostanza tossica per l'altra, e questo ad un tasso costante, indipendente dalla densitŕ dell'altra.
Ciň equivale ad introdurre i termini -ky e -jx rispettivamente nella prima e nella seconda equazione del sistema (2)
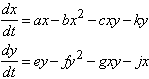
questo modello prevede l'estinzione della specie inizialmente meno abbondante e dunque poco aggiunge a quanto discusso.
|
Da quanto discusso, appare plausibile che l'origine dell'asimmetria biologica possa essere ricercata nella storia evolutiva della vita. L'approccio proposto permette anche di giustificare perché i D amminoacidi non siano del tutto assenti in natura. Infatti, ad esempio, nella capsula del carbonchio č contenuto l'acido D glutammico, e le D aminoacidossidasi, nonostante la scarsezza in natura di D amminoacidi sono piů diffuse ed abbondanti delle L aminoacidossidasi. Quest'ultimo č un esempio assai raro in natura dell'esistenza di enzimi per entrambe le forme stereoisomere dello stesso substrato: generalmente esistono enzimi per la sola forma L. Ciň premesso, se si considera che i D amminoacidi non partecipano mai al ciclo fondamentale acidi nucleici-proteine, possiamo spiegare la loro presenza ipotizzando che quando si verificň la competizione fra le due specie enantiomorfe sopravvissute, quella che disponeva casualmente di un ciclo riproduttivo piů efficiente oppure usufruiva di una maggior disponibilitŕ di substrato asimmetrico (L amminoacidi) orientň la sua evoluzione usufruendo di quei processi che ne permettevano la massima utilizzazione, lasciando che i D amminoacidi partecipassero occasionalmente a funzioni accessorie.
Un'altra prova che si aggiunge a favore dell'ipotesi di una selezione biologica č offerta dall'analisi della composizione dei meteoriti. Due meteoriti furono esaminati con garanzia di non contaminazione terrestre: il meteorite caduto vicino a Murray (Kentucky, 1950) e quello esploso sopra Murchison (Australia, 1969). dai frammenti del meteorite di Murchison vennero separate piccole quantitŕ di diciotto amminoacidi diversi. Di questi, sei sono presenti nelle proteine dei tessuti viventi, gli altri dodici ricorrono raramente o mai. Il meteorite di Murray diede risultati analoghi. |
Bibliografia:
V. Volterra - Variazioni e fluttuazioni del numero di individui in specie animali conviventi, Mem. accad. Naz. Lincei 1926
E. Havinga - Biochim. Biophys. Acta 1954
G. Wald - Annals N.Y. Acad. Sci. 1957
M. Calvin - Chemical evolution, Clarendon press, Oxford 1969
J. Maynard Smith - L'ecologia e i suoi modelli - Mondadori, EST 1975