2. Applicazione della spettroscopia di fluorescenza allo studio della struttura del nucleosoma
(2)
2a. Introduzione
Il nucleosoma è una complessa struttura macromolecolare, composta da DNA e proteine, che negli eucarioti rappresenta l’unità di base dell’organizzazione strutturale del patrimonio genetico. Esso fornisce un sistema per il necessario "impacchettamento" del DNA, che, pur essendo lungo milioni di coppie di basi, deve entrare nel nucleo della cellula. Inoltre il nucleosoma deve essere in grado, al momento opportuno, di rendere il DNA accessibile per la duplicazione e per la trascrizione. Si pensa che questo possa avvenire grazie a variazioni nella struttura del nucleosoma, quindi negli esperimenti presentati in questo articolo sono stati indotti cambiamenti conformazionali variando la forza ionica della soluzione, per poi misurare parametri relativi alla fluorescenza di sonde precedentemente legate ad esso.
La componente proteica del nucleosoma è costituita da due copie di quattro proteine (istoni), organizzate in un ottamero. In particolare l’istone H3 presenta un unico residuo di cisteina, a cui sono state legate le sonde iodoacetiletilendiammina-(1,5)-naftol solfonato (IAEDANS) e iodoacetilamminofluoresceina (IAAF). Il processo di labelling è stato condotto in soluzione di urea 5M e NaCl 2M, in queste condizioni la struttura del nucleosoma espone i residui di cisteina all’azione degli agenti chimici. Successivamente, tramite dialisi e successiva purificazione per centrifugazione, è stata ottenuta una soluzione di nucleosoma marcato, che è risultata indistinguibile da quello non marcato secondo i criteri di sedimentazione in gradiente di densità di saccarosio e digestione enzimatica, garantendo quindi che la denaturazione era di carattere reversibile.
2b. Intensità di fluorescenza
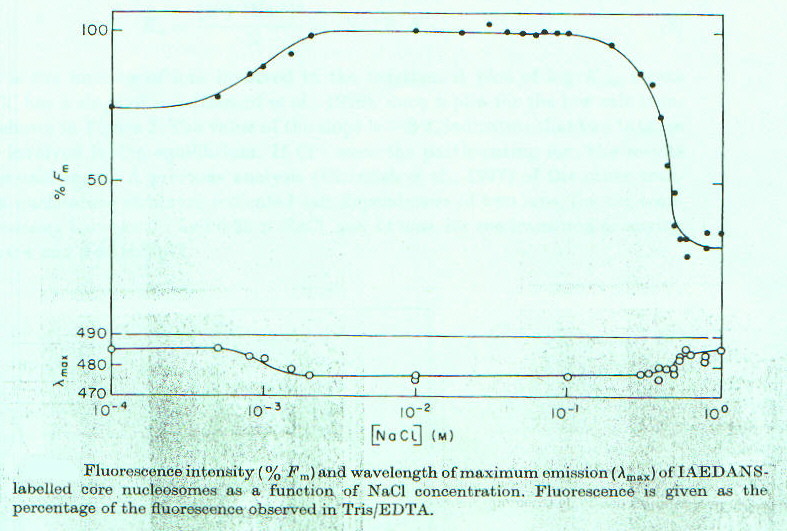
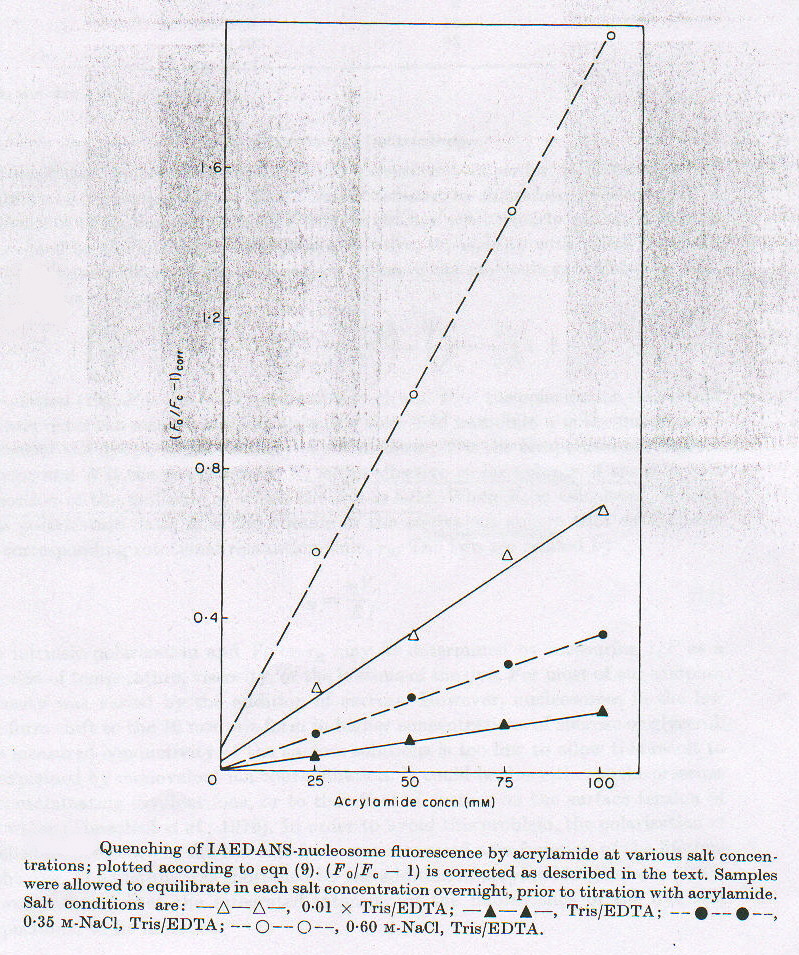
Utilizzando nucleosomi marcati con IAEDANS si nota una intensità di fluorescenza minore in condizioni di forza ionica particolarmente bassa e alta, rispetto a quanto avviene per valori intermedi, cioè nella soluzione tampone Tris/EDTA (a cui si associa il valore 100%, fig.2-1). E’ probabile che a forza ionica alta ci sia un indebolimento delle interazioni DNA-proteine, che sono principalmente di natura elettrostatica, mentre a forza ionica molto bassa sono sfavorite le interazioni idrofobiche, che sono alla base delle associazioni proteina-proteina. In entrambi i casi il complesso risulta avere una struttura meno compatta rispetto a quella che ha per valori intermedi di forza ionica, e questo si riflette sull’intensità di fluorescenza sia perché cambia l’abiente in cui la sonda si trova (da idrofobico a polare), sia perché quando la struttura è più aperta la sonda è più soggetta all’azione dei quencher presenti nella soluzione. Quest’ultimo fatto si può evidenziare aggiungendo concentrazioni crescenti di acrilammide (quencher) a soluzioni a diversa forza ionica di nucleosoma marcato. Come si può vedere in figura 2-2, l’azione del quencher è più efficace a valori molto alti (o ) o molto bassi (D ) di forza ionica, rispetto ai valori intermedi (in ordinata: F0/Fc – 1 , con F0 fluorescenza della sonda in assenza di quencher e Fc fluorescenza in presenza di quencher a concentrazione c).
2c. Polarizzazione di fluorescenza
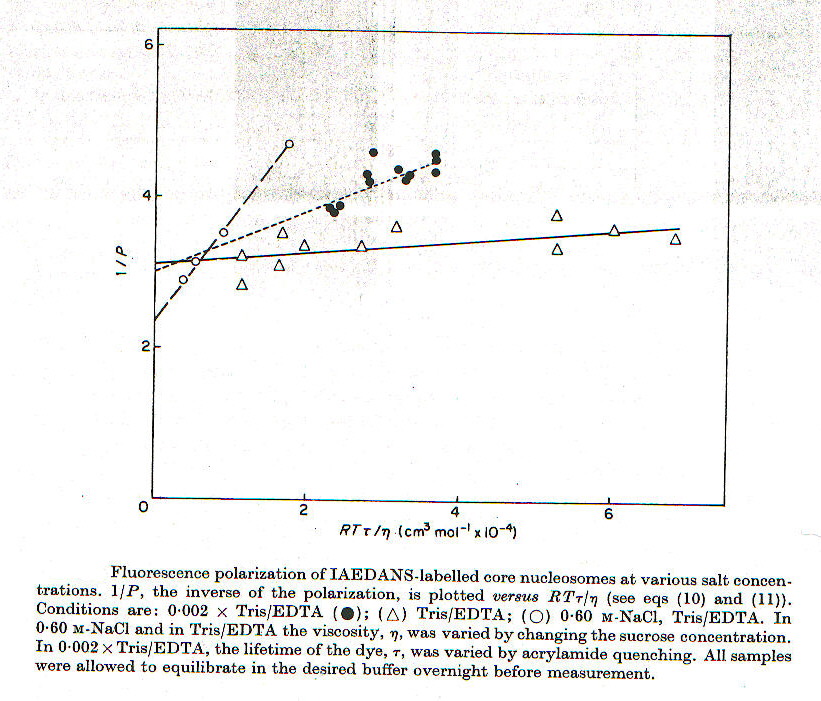
Utilizzando tecniche analoghe a quelle descritte nel paragrafo 1d, è stata misurata la polarizzazione in funzione della viscosità della soluzione, variata mediante l’aggiunta di saccarosio, da cui è stato possibile ottenere i valori del volume idrato Vh e il tempo di correlazione rotazionale tc. I risultati ottenuti sono presentati nel grafico in figura 2-3 e in tabella 2-1. I valori molto piccoli sia di tempo che di volume ottenuti a forza ionica molto bassa e molto alta sono probabilmente dovuti a moti interni del nucleosoma, cioè a movimenti indipendenti della zona C-terminale dell’istone H3. I dati ottenuti con il tampone Tris/EDTA e con altre soluzione a forza ionica leggermente diversa (dati non riportati) si adattano meglio, in base a dati indipendenti da questi come peso molecolare e densità, ad un ellissoide piuttosto che ad una sfera.
2d. Sistema a trasferimento di energia
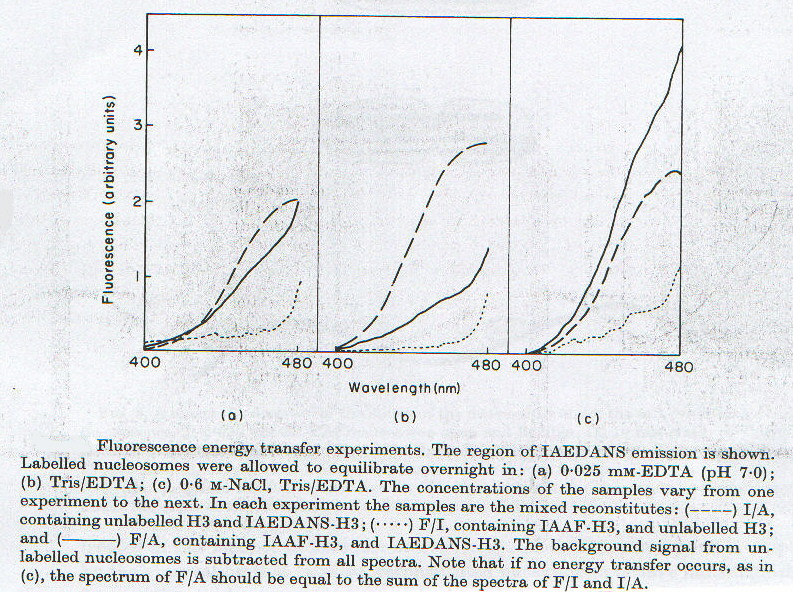
In questi esperimenti sono stati utilizzati quattro tipi diversi di nucleosomi marcati, ottenuti mescolando, prima della rinaturazione, soluzioni contenenti nucleosoma trattato con l’una o l’altra sonda, oppure nessuna delle due, in diverse percentuali. Così facendo i quattro campioni erano costituiti da nucleosomi marcati con entrambe le sonde, con una sola o con nessuna. La fluorescenza del campione non marcato è stata sottratta a tutte le altre e i diagrammi così ottenuti sono mostrati in figura 2-4; dalla fluorescenza del campione con entrambe le sonde è stata ancora sottratta quella del solo accettore (che comunque è molto bassa nell’intervallo di frequenze considerato), poi questa quantità è stata rapportata alla fluorescenza del solo donatore per ottenere E, secondo la formula:
E = 1 - [(FF/A - FF)/FA] = 1 - (fF/A/fA)
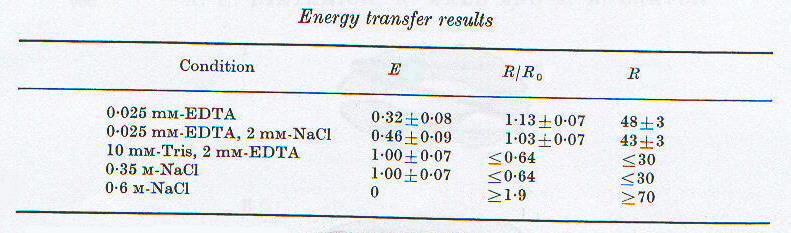
dove A = IAEDANS (donatore) e F = IAAF (accettore). La media di questi valori sulle diverse lunghezze d’onda, corretta per i necessari fattori stechiometrici in funzione della composizione della soluzione, porta ai valori di E riportati nella tabella 2-2. Da questi, noti i valori di R0 nelle diverse condizioni sperimentali, sono stati ricavati i valori di R. Anche questi dati, coerentemente con i precedenti, mostrano una distanza minima in condizioni intermedie, che poi aumenta a forza ionica minore e maggiore.
2e. Conclusioni
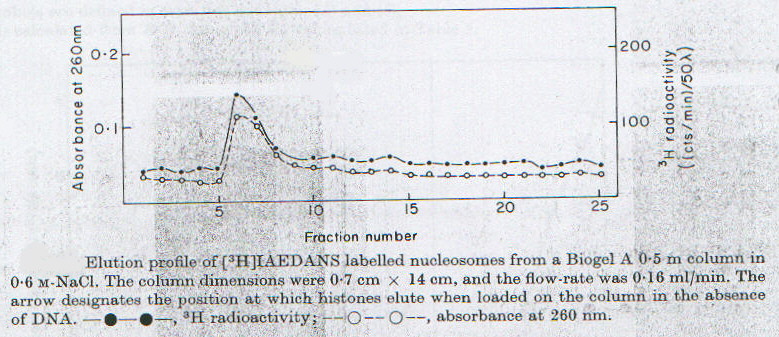
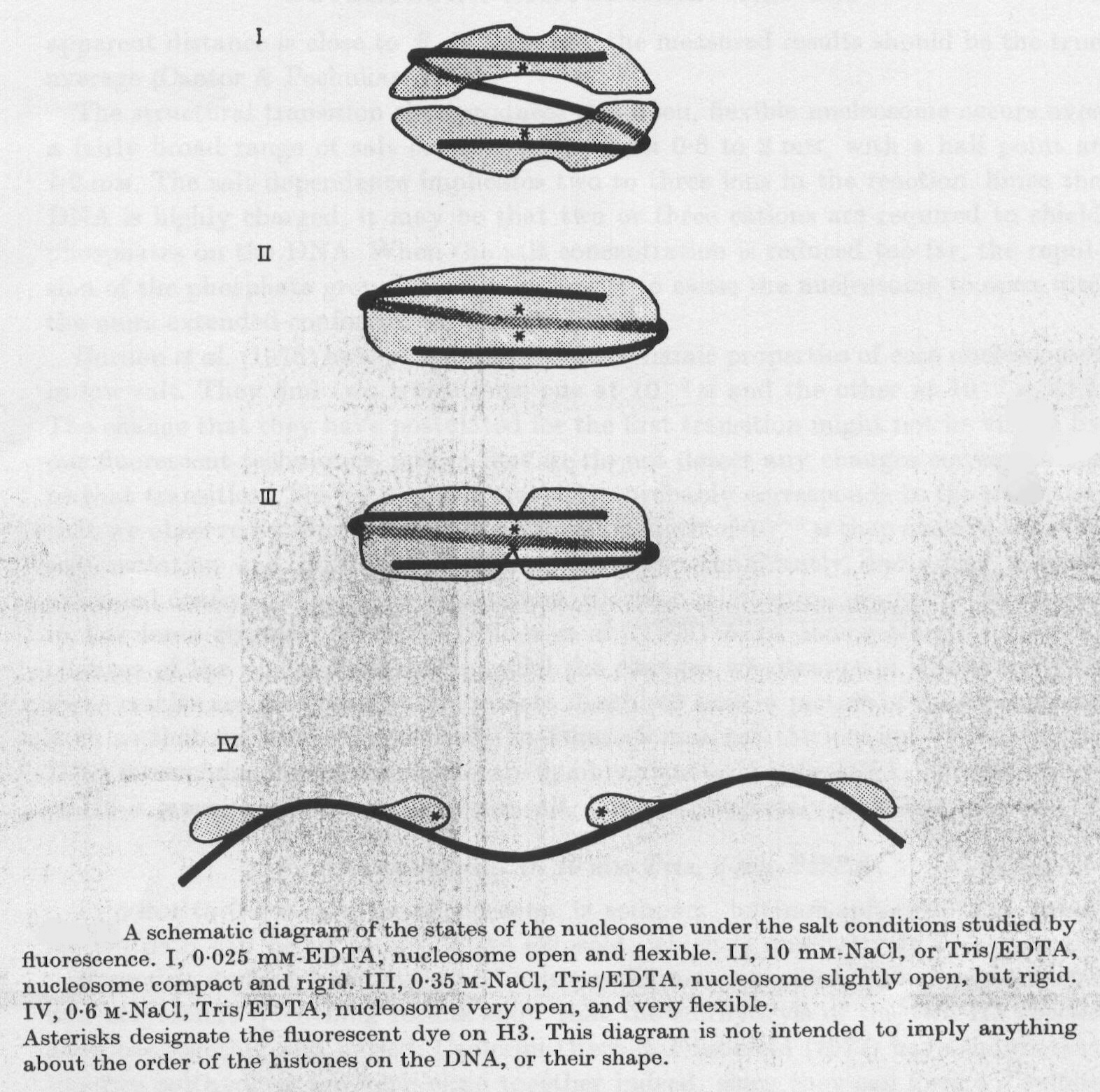
Per poter trarre le opportune conclusioni è necessario chiarire se a forza ionica elevata (0.6M NaCl) si ha dissociazione dell’istone H3 dal DNA oppure no. La risposta a questa domanda si trova nel grafico in figura 2-5, dove si dimostra che il segnale radioattivo dovuto alla sonda triziata si sovrappone perfettamente a quello di assorbimento a 260 nm, caratteristico del DNA, dopo gel filtrazione in colonna equilibrata con soluzione 0.6M di NaCl. Quindi anche in queste condizioni proteina e DNA non si separano, e pertanto le osservazioni sperimentali fin qui illustrate, unitamente a prove indipendenti fornite da microscopia elettronica e diffrazione di raggi X, portano al modello illustrato in figura 2-6. Tutto ciò dimostra la capacità del nucleosoma in vitro di mantenere diverse strutture e fornisce la condizione necessaria, ma non sufficiente, perché queste modificazioni conformazionali possano presentarsi anche in vivo, modulando lo stato funzionale di specifici segmenti del genoma.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}