3c. Struttura dell'intero ribosoma a 7.8A di risoluzione
(6)
Per ottenere la struttura dell'unità complessiva (70S) del ribosoma è stato necessario complessare le due subunità 30S e 50S con mRNA e tRNA o suoi analoghi (in particolare in questo lavoro sono stati utilizzati tRNAfMet interi, tRNALys deacilati e frammenti di tRNAPhe). Poiché questa struttura è stata ottenuta a risoluzione inferiore rispetto alle precedenti, gli aspetti più interessanti e nuovi che si possono ricavare da questo lavoro, rispetto a quelli precedentemente descritti, consistono nella possibilità di avere una visione di insieme dell'intera struttura ribosomiale, quella cioè che in vivo opera effettivamente la sintesi proteica, e nello studio delle interazioni delle due subunità tra di loro e con le molecole di tRNA.
In figura 3c-1 è riportata una mappa di densità elettronica, in cui la parte scura rappresenta la subunità 30S e la parte chiara quella 50S. E' possibile confrontare questa struttura con quelle delle singole subunità riportate nelle figure 3a-1 e 3b-1, notando la perfetta coincidenza dei vari domini e delle diverse proteine presenti (H, S, P, B, CP, L1).
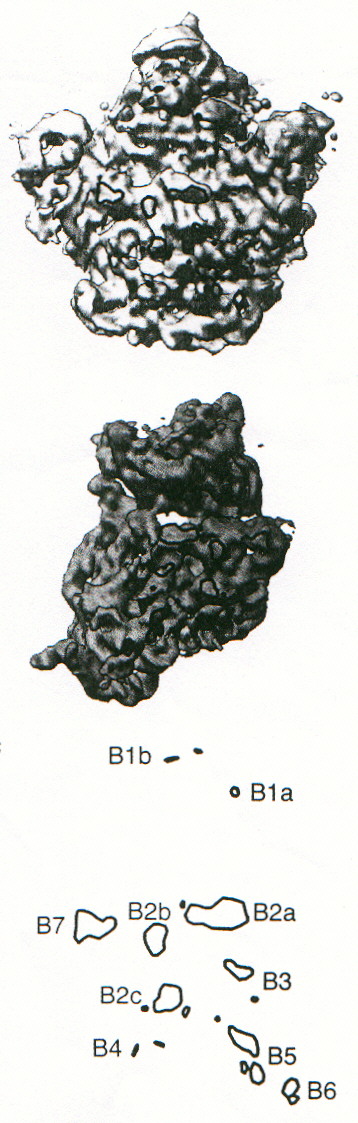
In figura 3c-2 sono stati messi in evidenza i punti di contatto tra le due subunità. Questi "ponti" (B = bridge) possono essere costituiti da interazioni dirette tra le molecole di RNA (soprattutto a doppia elica) presenti nelle due subunità, o possono richiedere la mediazione di proteine di una delle due subunità, che interagiscono con i solchi di doppia elica di RNA presenti sull'altra.
Per mezzo dei tre complessi, citati all'inizio del paragrafo, con tRNA e analoghi, confrontando mappe di densità elettronica e dati di microscopia crioelettronica, è stato possibile individuare i tre siti di legame dei tRNA con il ribosoma: i siti A, P ed E (= exit). Da notare, in figura 3c-3, la posizione pressoché parallela dei siti P e A, fondamentale per la formazione del legame peptidico, mentre il sito E diverge da questa direzione. Un'analisi più dettagliata dei punti di interazione tra i vari tRNA e il ribosoma ha messo in evidenza dei dati strutturali in accordo con la funzione biologica che i diversi siti devono assolvere. Ad esempio il tRNA che si trova nel sito P risulta essere fortemente legato in più punti al ribosoma e in maniera sostanzialmente sequenza indipendente, rispettando l'esigenza principale di trattenere la catena peptidica nascente, indipendentemente dal tRNA. La selezione del tRNA corretto, fondamentale per la corretta traduzione dell'informazione genetica, avviene invece nel sito A dove il corretto appaiamento codone-anticodone è essenziale per l'interazione con il tRNA che trasporta l'amminoacido da aggiungere. Infatti in assenza di tRNA l'mRNA è praticamente non legato al ribosoma, mostrando una struttura flessibile e aperta necessaria per consentire l'ingresso del tRNA legato al fattore EF-Tu, ma anche per permettere l'assestamento del tRNA dopo il rilascio di tale fattore, e la formazione di una struttura ordinata, solo a patto che ci sia perfetta coincidenza tra codone e anticodone. In figura 3c-4 è riportato un modello in cui i tre siti sono sovrapposti alla mappa di densità elettronica della subunità 30S del ribosoma.
La coincidenza di alcuni punti di contatto tra le due subunità con punti di interazione tra tRNA e ribosoma, unitamente a prove indipendenti ottenute con l'uso di antibiotici che bloccano la sintesi proteica, suggerisce una connessione funzionale tra l'associazione delle due subunità e il legame con tRNA. Questa connessione è anche attuata tramite i movimenti che dati recenti sembrano suggerire coinvolgere l'intero ribosoma durante il suo funzionamento.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}