3a. La struttura della subunità 30S alla risoluzione di 5.5A
(4)
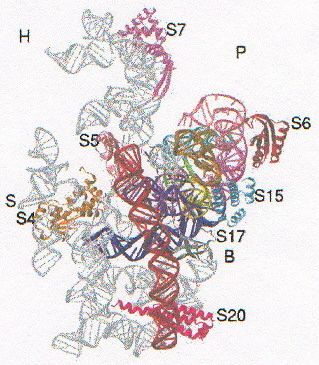
La mappa elettronica della subunità ribosomiale 30S a 5.5A di risoluzione ha permesso innanzitutto di confermare (anche quantitativamente) le strutture già note da studi di microscopia elettronica (fig. 3a-1, la faccia mostrata è quella di interazione con la subunità 50S): testa (H = head), piattaforma (P), spalla (S) e corpo (B = body).
In questa mappa è stato possibile individuare chiaramente le regioni di RNA a doppia elica (in figura 3a-2 le due linee rosse rappresentano una struttura a doppia elica di tipo A sovrapposta alla densità elettronica), le proteine a struttura nota, le regioni di interazione tra RNA a doppia elica e proteine (fig. 3a-3, in rosso la proteina S6 e in azzurro la S15), nonché tentare di individuare la struttura e la posizione di proteine non ancora note. Tutto questo è stato possibile grazie al fatto che nella mappa elettronica a questa risoluzione le a-eliche appaiono come tubi, mentre le regioni b-sheet e i loop sono a densità elettronica inferiore.
In particolare è stato analizzato il dominio centrale della molecola di RNA 16S presente in questa subunità ribosomiale, costituito da un insieme compatto di doppie eliche di RNA, in cui un'elica si insinua nel solco minore di una doppia elica adiacente (fig. 3a-4): esso contiene molti importanti siti di interazione con varie proteine e comprende buona parte della piattaforma, con i suoi siti essenziali per l'associazione delle subunità e per il legame con il tRNA del sito P. Ad un'estremità di questo dominio centrale è stata anche individuata una importante regione, H27, per cui è stato proposto un ruolo rilevante nell'elevato livello di accuratezza della traduzione. L'azione di H27 consisterebbe nella modulazione conformazionale del sito decodificante (A) per interazione diretta con la regione H44, che si trova subito sotto il sito A (fig. 3a-5), a dimostrazione del fatto che, lungi dall'essere una struttura rigida, come per molto tempo si è erroneamente ritenuto, il ribosoma deve molte delle sue proprietà catalitiche e di controllo a variazioni conformazionali sia delle componenti proteiche che di quelle ribonucleiche. Un altro esempio di questo è il fatto che il legame alternativo con IF3 o con la subunità 50S avviene in seguito a variazioni conformazionali all'interno della piattaforma.
Sebbene molte interazioni fondamentali per la struttura e il funzionamento del ribosoma avvengano a livello di interazione RNA-proteine, esiste almeno un esempio di interazione fondamentale di tipo proteina-proteina (fig. 3a-6).
In figura 3a-7 è riportato il modello complessivo, visto frontalmente, della subunità 30S del ribosoma, ricavato dai dati sperimentali riportati nell'articolo, con tutti i tratti di doppia elica di RNA individuati e le proteine note. L'aspetto più interessante che emerge da questo modello, soprattutto nella visione laterale in figura 3a-8, è l'assenza quasi completa di proteine all'interfaccia tra le due subunità ribosomiali, che suggerisce l'esistenza di un ipotetico ribosoma primordiale privo di proteine, anche se, come già illustrato, molte funzioni essenziali dei moderni ribosomi sono associate con specifiche proteine.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}