2c. Importanza del legame con ioni rame per la struttura dei prioni
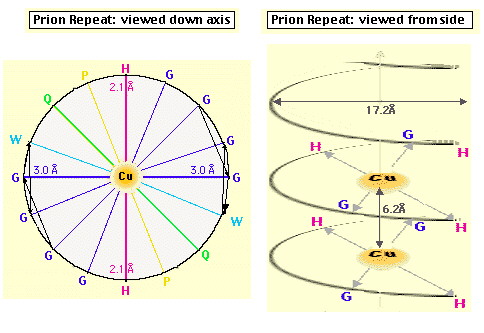
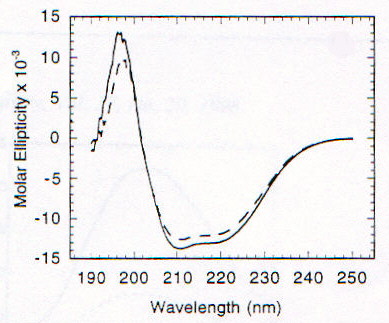
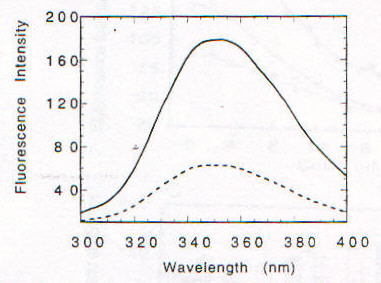
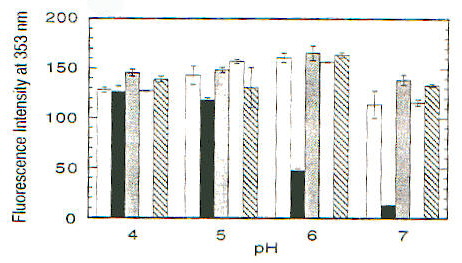
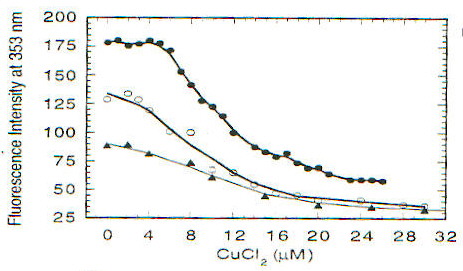
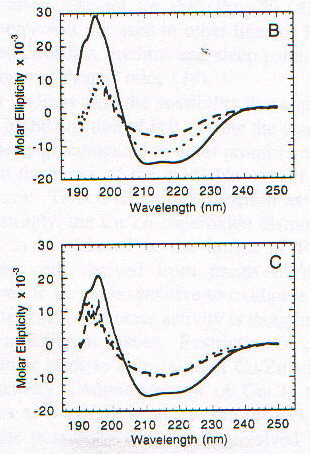
La funzione della proteina prionica nella forma PrPC non è chiara. Topi knock-out per la proteina prionica (cioè topi in cui sono state inattivate entrambe le copie del gene codificante per tale proteina), che non sviluppano la malattia se viene loro iniettato l’estratto di cervello di topi malati [1], crescono e vivono piuttosto normalmente, almeno per le prime 70 settimane (su circa 100) di vita; in realtà fenomeni di morte prematura sono stati osservati, ma il significato non è chiaro [13]. PrPC viene sintetizzata nel reticolo endoplasmatico, modificata nell’apparato del Golgi e trasportata sulla superficie cellulare, a cui rimane legata tramite un’ancora di glicofosfatidil inositolo (GPI) [2]; durante questo percorso PrPC viene glicosilata su due residui di Asp [15]. La sequenza amminoacidica della proteina prionica comprende circa 250 amminoacidi ed è altamente conservata in molte specie, nella forma matura PrPC (di topo) comprende i residui 23-231, di cui i primi 120 risultano avere una strutura disordinata in esperimenti NMR, in condizioni di assenza di ioni bivalenti [13]. Recentemente è stato scoperto che questa regione contiene un gruppo di ottameri (Pro-His-Gly-Gly-Gly-Trp-Gly-Gln) che sembrano legare ioni Cu(II), quindi una possibile funzione di PrPC potrebbe essere quella di fare da "shuttle" o da regolatore per ioni Cu2+, tra le altre cose fondamentale per l’attività di un ampio gruppo di enzimi ossidativi, importanti anche per evitare la distruzione di neurotrasmettitori (questo potrebbe spiegare l’importanza di PrPC nelle giunzioni sinaptiche) [13]. Da un punto di vista strutturale in particolare è stata messa in evidenza la possibilità che il legame con Cu2+ sia in grado di indurre una conversione della regione "disordinata" in a-eliche, stimolando poi la formazione di a-eliche nel resto della proteina, cosicché la sua mancanza potrebbe giocare un ruolo importante nello trasformazione maligna della proteina prionica [13]. In realtà si pensa che le strutture formate siano delle particolari strutture elicoidali (con diametro di 17.2A) in cui residui di His e Gly (con il gruppo C=O) si trovano disposti in modo tale da poter coordinare Cu2+ (fig. 2-20) [14]. Esperimenti condotti utilizzando ancora una volta il dicroismo circolare su una proteina prionica ricombinante di criceto indicata come ShaPrP(29-231), che risulta avere una conformazione molto simile a PrPC in base agli spettri CD, hanno portato alla conferma della capacità di Cu2+ di indurre variazioni conformazionali nella proteina [15]. Gli spettri riportati nelle figure 2-21 e 2-22, rispettivamente CD e fluorescenza (l di eccitazione = 280nm), mostrano infatti una notevole differenza tra la proteina in assenza di Cu2+ (linea continua) e in sua presenza (linea tratteggiata). Il legame con Cu2+ risulta inoltre essere specifico rispetto ad altri metalli, dipendente dal pH (fig. 2-23, le cinque barre rappresentano da sinistra a destra: proteina senza metalli, proteina con Cu2+, Zn2+, Co2+ e Mn2+) e saturabile: in figura 2-24 sono riportati i dati ottenuti per l’intera proteina (pallini neri), e per due segmenti (57-91, pallini bianchi, e 73-91, triangolini); in particolare poiché il segmento (73-91) è costituito da due soli ottapeptidi si conclude, in accordo con quanto visto prima, che sono necessarie due sequenze ripetute per legare uno ione rame. Quando però, sfruttando la capacità di ShaPrP(29-231) di mostrare una transizione non reversibile a-elica à b-sheet per riscaldamento, si nota che la presenza di Cu2+ fa aumentare la tendenza a formare b-sheet, sorge un disaccordo con il modello precedente. In figura 2-25 sono riportati gli spettri della proteina con (C) e senza (B) ioni rame, registrati secondo la sequenza di temperatura: 25°C (linea continua), 76°C (linea tratteggiata), 25°C (linea puntata); si può notare l’aumentata stabilità della struttura b-sheet dopo raffreddamento in presenza di ioni rame. Tutto questo porterebbe a concludere che il legame con il rame favorisce la formazione di PrPSc e quindi lo sviluppo della malattia, ma l’interpretazione degli spettri CD potrebbe essere forviata dalla presenza di ipotetiche strutture, come quella illustrata prima, non considerate quando si tenta di eseguire il fit della curva sperimentale.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}