2b. Studio della struttura dei prioni tramite dicroismo circolare e altre tecniche spettroscopiche
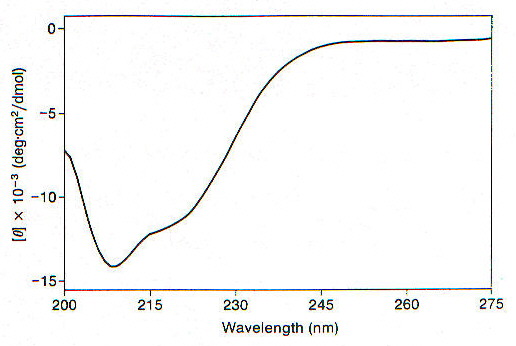
Il dicroismo circolare è stato applicato allo studio della struttura secondaria di PrPC [5]. Lo spettro CD ottenuto (fig. 2-3) mostra un minimo a 208nm e una spalla a 222nm, indicando la presenza di struttura a-elica, che il fit eseguito secondo quanto illustrato nel paragrafo precedente quantifica in 36%. I risultati ottenuti con la spettroscopia a raggi infrarossi con trasformata di Fourier (FTIR) sono simili (42% di a-elica, 3% di b-sheet – praticamente niente se si considerano i margini di errore), mentre mostrano una struttura ad alta percentuale di b-sheet (43% di b-sheet e 30% di a-elica) per PrPSc e una percentuale di b-sheet ancora più alta (54%) per il segmento PrP27-30. Quest’ultimo è un frammento N-troncato e proteasi-resistente di PrPSc (è da notare come lo stesso trattamento proteolitico eseguito su PrPC porta ad una degradazione completa della proteina), che costituisce l’unità infettiva minima della proteina prionica, comprende i residui 90-231 [10] ed è in grado di formare amiloidi a forma di bacchetta, mentre né PrPSc né PrPC formano aggregati di questo tipo, al limite possono formare strutture amorfe se seccate (studi di microscopia elettronica). Anche calcoli computazionali hanno dimostrato l’esistenza, all’interno della proteina prionica, di una struttura con notevole stabilità se disposta a quattro a-eliche, che però è pure molto stabile in conformazione b-sheet; anche studi di diffrazione di raggi X hanno convalidato il modello della conversione a-elica à b-sheet [6]. Inoltre poiché il blocco della sintesi proteica porta ad un arresto della formazione di PrPSc è probabile che nella transizione PrPC (a-elica) à PrPSc (b-sheet) sia essenziale l’azione di altre proteine cellulari, molto probabilmente tipo chaperonine.
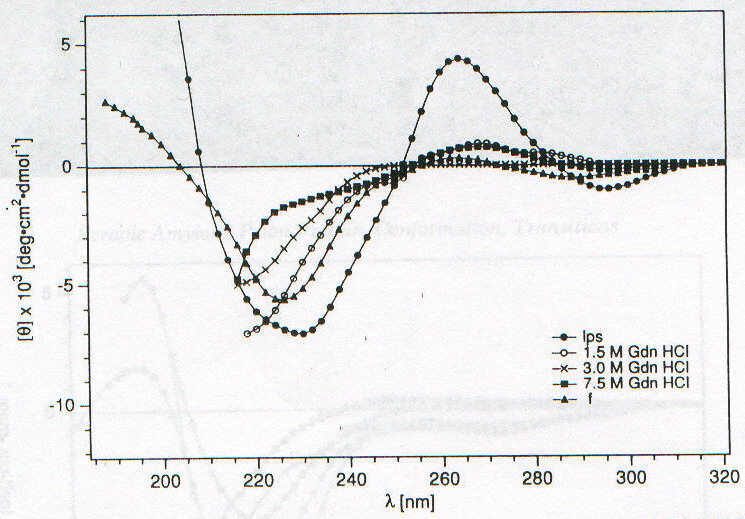
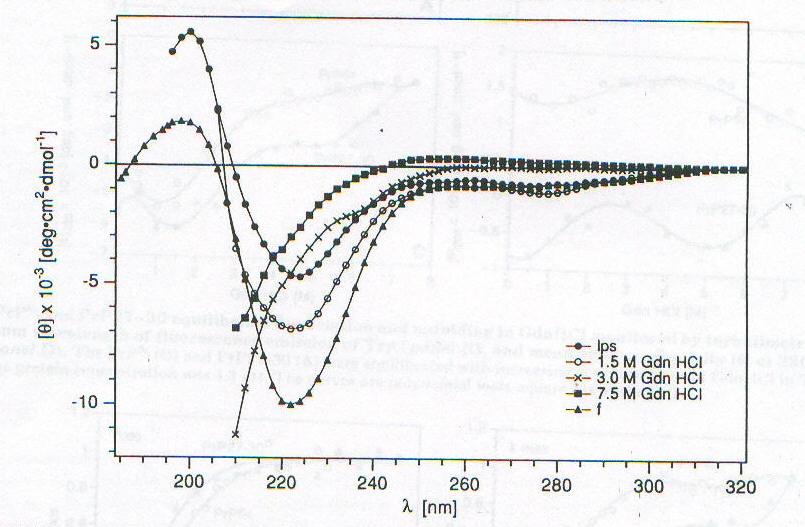
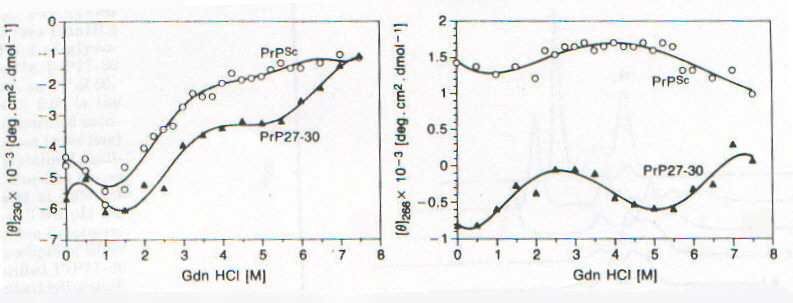
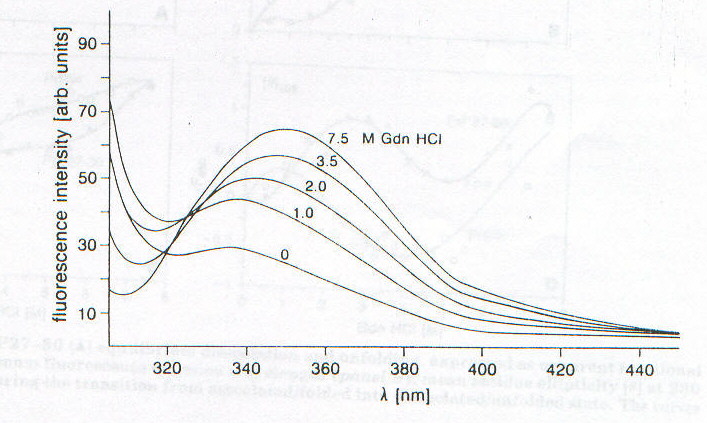
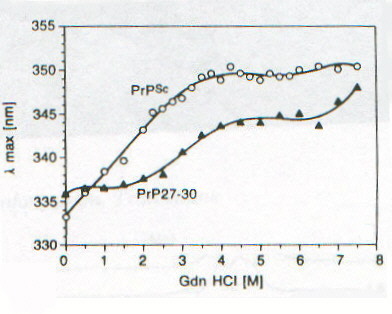
Lo studio di PrPSc e di PrP27-30 tramite CD è fortemente ostacolato dall’insolubilità che caratterizza queste due forme della proteina prionica; è stato infatti possibile ottenere spettri CD per queste due forme solo associate a liposomi o depositate come film sottili [7]. Nelle figure 2-4 e 2-5 sono riportati gli spettri CD di PrPSc e PrP27-30 rispettivamente: dopo dissociazione e incorporazione in liposomi (lps), dopo 24 ore di incubazione con diverse concentrazioni di guanidina in HCl (che provoca dissociazione e unfolding) e associata in stato solido in film sottile (f). Il fit eseguito secondo quanto riportato nel paragrafo precedente sulle due curve corrispondenti alle forme associate ai liposomi ha portato ai risultati riassunti nella tabella 2-1. L’uso dei liposomi ha lo scopo di superare la difficoltà dovuta all’insolubilità di queste proteine, mentre l’uso di solventi in grado di solubilizzarle porterebbe a denaturazione. I dati ottenuti mostrano un aumento della percentuale di b-sheet nel passaggio da PrPSc a PrP27-30 e un aumento ancora maggiore quando PrP27-30 forma aggregati amiloidi nello stato solido; anche se la probabile interazione diretta con la membrana del liposoma (suggerita dalla scomparsa dell’intensa banda intorno a 260nm dovuta probabilmente a Trp) dovrebbe indurre a interpretare con cautela i dati ottenuti. D’altronde la diminuzione della "banda aromatica" indotta sia dall’aggregazione che dalla guanidina è indice di processi di unfolding. Questi dati forniscono una prova diretta dell’importanza della struttura b-sheet nel processo di formazione delle placche amioloidi e della loro capacità di stimolare le transizioni necessarie per la formazione delle stesse placche. La mancanza di un punto isodicroico (unico punto di intersezione tra le varie curve) indica la presenza di più intermedi con diversa stabilità, dovuti probabilmente a domini con differente stabilità conformazionale. Un processo più complesso di quello a due stadi è suggerito anche dalla complessità dell’andamento degli spettri CD rispetto alla concentrazione di guanidina, apprezzabile ancor meglio nelle curve di dissociazione e unfolding riportate in figura 2-6. La spettroscopia di fluorescenza (fig. 2-7, per PrPSc) fornisce dei risultati per alcuni aspetti forvianti, in quanto mentre l’aumento di lmax con la concentrazione di guanidina è compatibile con i prevedibili processi di dissociazione/ unfolding che espongono i residui di Trp al solvente, l’aumento di intensità di emissione, che suggerirebbe esattamente il contrario, è dovuta a fenomeni di light scattering; è per questo che per seguire tali processi si utilizza lmax e non il valore dell’intensità di fluorescenza (fig. 2-8). I dati sperimentali sono compatibili con un modello a quattro step: aggregati à monomeri dissociati folded à intermedi parzialmente unfolded à monomeri unfolded.
Lo studio di polipeptidi più o meno lunghi, modificati tramite tecniche di biologia molecolare o addirittura sintetizzati chimicamente, piuttosto che dell’intera proteina PrPSc, fornisce un utile strumento di indagine, nonché un modello più semplice da analizzare. Spesso così facendo si risolvono infatti i problemi di solubilizzazione della proteina; inoltre poiché molti scienziati ritengono che nell’evoluzione delle patologie legate ai prioni sia di fondamentale importanza la loro parziale proteolisi, lo studio di singoli segmenti delle proteine prioniche potrebbe avere un significato biologico più immediato di quanto non si possa pensare. Il problema è che, non essendoci un comune accordo nella scelta della parte essenziale della proteina per l’attività biologica dei prioni e per lo sviluppo della malattia, diversi gruppi di ricercatori hanno usato criteri diversi (conservazione della sequenza in specie diverse, siti del taglio proteolitico) nel fare questa scelta, e di conseguenza hanno concentrato la loro attenzione su diversi peptidi, ottenendo risultati a volte contrastanti e proponendo meccanismi molecolari differenti.
Un modello per la formazione di placche amiloidi è stato trovato in un oligopeptide sintetico costituito da sedici amminoacidi [8]. Studiando infatti peptidi a composizione diversa, ma con la caratteristica di essere costituiti da residui carichi alternati a residui idrofobici (alanina) disposti in modo tale da autocomplementarsi stabilizzando una struttura b-sheet, uno in particolare ha mostrato un comportamento insolito. La sequenza di questo peptide (indicato come DAR16-IV) è:
- - - - + + + +
Ac-NH-ADADADADARARARAR-CONH2
In
figura 2-9 sono riportati gli spettri CD di questo peptide in condizioni di diversa temperatura. Nello spettro A si vede che il riscaldamento (10min a 90°C, riportato poi a 20°C) porta ad una transizione strutturale da b-sheet direttamente ad a-elica, senza un intermedio random-coil osservabile; in B i numeri indicano i minuti di riscaldamento a 75°C (lo spettro è stato registrato dopo raffreddamento a 20°C): il processo richiede un riscaldamento di almeno 8 minuti e oltre 32 non si osservano ulteriori variazioni; l’a-elica che così si forma è piuttosto stabile: in C i numeri indicano la temperatura a cui sono stati registrati gli spettri (a-elica passa da 60% a 30% andando da 10°C a 90°C), inoltre l’a-elica è stabile per settimane a temperatura ambiente. Nello stesso lavoro si possono trovare esperimenti in cui vengono variati la concentrazione del peptide, la forza ionica della soluzione e il pH. Considerando che questi polipeptidi nella forma b-sheet danno origine spontaneamente ad aggregati proteici macroscopici, la cui rottura richiede un certo tempo (si noti la necessità di almeno 8 minuti di incubazione), questi esperimenti forniscono un modello per la formazione di placche amiloidi, in quanto sono prove sperimentali di una possibile transizione a-elica à b-sheet, cineticamente molto lenta, che però una volta avvenuta porta alla formazione di placche insolubili molto stabili.Lo studio del segmento C-terminale della proteina prionica cellulare di topo PrP(121-231)
[9] è stato condotto utilizzando cloruro di guanidinio (GdmCl) come agente denaturante e seguendo il processo tramite dicroismo circolare. Lo spettro CD riportato in figura 2-10 mostra la quasi perfetta sovrapposizione delle curve misurate prima dell’aggiunta di GdmCl (linea continua) e dopo la sua aggiunta e successiva dialisi (linea puntata), mentre lo spettro registrato in presenza di GdmCl (linea tratteggiata) rivela una struttura unfolded. La reversibilità e la natura cooperativa del processo di unfolding/refolding, nonché il tipico aspetto di un processo a due stadi, sono evidenti nel grafico in figura 2-11, in cui sono riportati i valori di [q] a 222nm in funzione della concentrazione di GdmCl per la proteina nativa (pallini bianchi, processo di unfolding) e per quella precedentemente denaturata (pallini neri, processo di refolding). L’energia libera di folding ricavabile da questi dati è di –21.8 (± 1.4) kJ/mol. Il modello strutturale che emerge è in disaccordo con la struttura di PrPC proposta da modelli molecolari secondo cui dovrebbero esserci quattro a-eliche, in quanto manca la prima elica.Un altro studio sul dominio PrP(121-231) è stato condotto inducendo l’unfoding di questo dominio diminuendo il pH e per trattamento con urea
[10]; lo scopo di questi studi è quello di determinare la possibilità che anche questo dominio C-terminale della proteina prionica sia in grado di subire cambiamenti conformazionali, essendo ormai noto che il peptide 90-120 (disordinato in PrPC e resistente alla proteasi in PrPSc) costituisce la minima differenza strutturale tra le due forme della proteina prionica. Mentre gli esperimenti precedentemente descritti, condotti a pH 7, hanno mostrato un processo a due soli step, la denaturazione con urea a pH bassi mostra un intermedio composto principalmente di b-sheet, d’altronde a pH 7 anche in questo caso non si hanno intermedi (fig. 2-12); in tutti i casi il processo risulta reversibile. In figura 2-13 è riportato il processo di denaturazione a concentrazione costante di urea (3.5 M) in funzione del pH. Attraverso la registrazione degli spettri CD delle tre forme della proteina (nativa, linea continua; intermedio, linea puntata; denaturata, linea tratteggiata; fig. 2-14) è stato possibile seguire l’andamento delle frazioni delle tre specie, secondo quanto illustrato in figura 2-15 (D = proteina Nativa, = proteina Unfolded, pallini neri = Intermedio). Bisogna anche considerare che normalmente gli intermedi presenti nei processi di denaturazione delle proteine assomigliano nella loro struttura secondaria alla proteina nativa, mentre qui questo non accade. Tutti questi dati suggeriscono quindi lo schema illustrato in figura 2-16, secondo cui il pH acido degli endosomi, in cui PrPC, normalmente esposta sulla superficie di membrana a un pH fisiolgico, si potrebbe trovare in seguito a endocitosi, porterebbe alla formazione reversibile dell’intermedio b-sheet, che però potrebbe aggregarsi irreversibilmente per dare oligomeri di PrPSc.Uno degli autori di questo lavoro ha poi firmato, in tempi più recenti, un altro articolo
[11] in cui si evidenzia non solo la mancanza di intermedi nel processo di denaturazione di PrPC a pH 7, ma anche l’estrema rapidità di questo processo. Al fine di studiare l’aspetto cinetico di questa trasformazione si è utilizzato il solo dominio PrP(121-231) della proteina prionica cellulare di topo, mutata in modo tale da sostituire Phe 175 con Trp 175. In questo modo, pur ottenendo lo stesso spettro CD e la stessa curva di unfolding/refolding seguita tramite CD (fig. 2-17), e avendo quindi la garanzia che la mutazione puntiforme non ha portato variazioni significative né nella conformazione né nell’andamento del processo, si ha un segnale di fluorescenza fortemente dipendente dal grado di unfolding della proteina. Perciò è stato possibile misurare l’intensità di fluorescenza nel tempo utilizzando la tecnica stopped flow (a 4°C perché altrimenti il processo sarebbe stato troppo veloce) ottenendo i grafici riportati in figura 2-18. In tutti i casi i dati sperimentali si adattano molto bene a un andamento monoesponenziale con costanti di tempo pari a 2.33ms (grafico a: concentrazione di urea 3.7M) e a 9.62ms (grafico b: concentrazione di urea 6.5M; 1 = refolding, 2 = unfolding, da notare l’identico andamento per i due processi). In conclusione il processo di unfolding/refolding risulta reversibile, molto veloce (uno dei più veloci misurati finora) e a due step, perciò il modello precedentemente descritto (in cui cioè un intermedio cinetico di folding di PrPC porta alla formazione di PrPSc) risulta improbabile a pH 7, pur rimanendo possibile che a pH più basso lo stato intermedio unfolded potrebbe essere un precursore di PrPSc, a patto che quest’ultima sia in grado di interagire con tale intermedio.Un esempio della diversità di risultati che si possono ottenere studiando parti diverse della proteina prionica si può trovare in un lavoro
[12] in cui l’importanza del "cuore" idrofobico della proteina prionica, presente all’interno del peptide PrP(106-126), per la sua attività neurotossica è stata studiata sostituendo, tramite tecniche di biologia molecolare, alcuni residui idrofobici di questa sequenza con residui di serina, a carattere idrofilico. I test di neurotossicità, eseguiti su neuroni di topo in coltura, hanno dimostrato come la diminuzione di idrofobicità è parallela alla diminuzione di neurotossicità del peptide, mentre tramite dicroismo circolare è stato possibile evidenziare che PrP(106-126) è caratterizzato da un passaggio da random coil (pH 5, minimo a 195nm) ad alta percentuale di b-sheet (pH 8, minimo intorno a 220nm) con conseguente capacità di formare fibrille amiloidi; è da notare come questo avviene invece a pH acido per la porzione di proteina PrP(121-231). Infatti le interazioni idrofobiche portano i peptidi vicini e facilitano la formazione di strutture b-sheet e di aggregati, mentre la sostituzione con residui idrofilici rompe queste interazioni. I peptidi mutanti mantengono invece tutti una certa percentuale di random coil (fig. 2-19, la linea sottile porta per confronto lo spettro CD della proteina non mutata) e solo i peptidi che hanno una certa percentuale di b-sheet e una minor quantità di random coil (A/-, sesta riga) sono in grado di provocare degenerazione dei neuroni in coltura. Comunque non tutti i peptidi che formano b-sheet sono necessariamente neurotossici (ad esempio AAAA/SSSS, quarta riga), indicando che ci potrebbero esssere altri fattori importanti nel determinare la neurotossicità.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}