2. Studi conformazionali sui prioni
2a. Il dicroismo circolare (CD) nello studio della struttura secondaria delle proteine
[4]Il dicroismo circolare fa parte delle spettroscopie chiroottiche, cioè di quelle tecniche spettroscopiche che, utilizzando luce polarizzata, mettono in risalto l’attività ottica delle molecole in esame. Per attività ottica si intende appunto la capacità di una molecola di ruotare il piano di polarizzazione della luce incidente. Tale proprietà deriva da una asimmetria della molecola che porti al fatto che la sua immagine speculare non sia ad essa sovrapponibile (chiralità; le due strutture speculari non sovrapponibili si definiscono enantiomeri). Condizione sufficiente, ma non necessaria, affinché una molecola sia chirale è la presenza di un atomo di carbonio asimmetrico (cioè con quattro sostituenti diversi), e questo è fondamentale nei processi di riconoscimento di molti composti biologicamente attivi. La possibilità di studiare la struttura secondaria delle proteine (e anche degli acidi nucleici) con le spettroscopie chiroottiche si basa sul fatto che particolari strutture, come le eliche sinistrorse e destrorse, hanno una propria attività ottica, oltre a quella degli atomi di carbonio asimmetrici, presenti ad esempio nei singoli amminoacidi. In particolare nel dicroismo circolare la radiazione incidente è costituita da due componenti di uguale ampiezza E e di polarizzazione circolare opposta, ciascuna a sua volta composta da due onde piano-polarizzate, secondo le equazioni:
ER = ½ (iE0sin
wt + jE0coswt)EL = ½ (iE0sin
wt - jE0coswt)dove i caratteri in neretto indicano grandezze vettoriali e i
^ j. Poiché le due componenti ER e EL sono chirali e tra di loro "enantiomeri", una molecola chirale interagisce diversamente con ciascuna di esse sia in termini di deviazione del piano di polarizzazione, sia in termini di differente assorbimento (fig. 2-1). In partcolare in CD si considera la differenza di assorbanza rispetto alle due componenti (AL e AR) e si definisce l’ellitticità q (in gradi) come:q
= 2.303(AL - AR)180/4pe normalizzando rispetto alla concentrazione (moli per litro) del campione (C) e al cammino ottico (l) si ottiene l’ellitticità molare:
[
q] = 100q/Cl = 3300Dedove
De = eL – eR, ricordando la legge di Beer-Lambert A = elC. Il CD è dunque una spettroscopia di assorbimento e quindi la condizione necessaria, ma non sufficiente, per avere un segale CD in un certo intervallo di frequenze è quella di avere assorbimento in quello stesso intervallo (me¹ 0). In più per avere segnale CD devono essere verificate le condizioni m¹ 0 e me non perpendicolare a m, dove me è il momento di dipolo elettrico di transizione e m è l’analogo momento magnetico. Infatti si definisce la forza rotazionale R:R = (3hc/8p3N0)¦ { [ q ( l ) ] / l } dl = imag½ me.m ½
dove h è la costante di Planck, N0 il numero di Avogadro e c la velocità della luce nel vuoto.
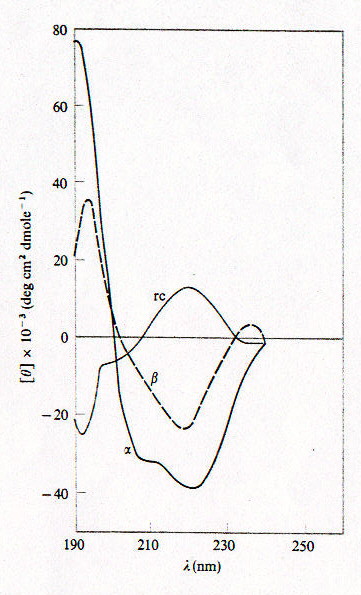
Gli spettri CD si ottengono graficando [q] in funzione di l (che per le studio di proteine è generalmente nella banda dell’ultravioletto). Poiché gli spettri (fig.2-2) corrispondenti alle strutture secondarie a-elica (a), b-sheet (b) e random coil (rc), ottenuti da spettri di proteine la cui struttura era già nota con altri metodi, sono piuttosto diversi, è possibile ricavare, entro un certo margine di errore, le percentuali delle tre strutture secondarie dallo spettro CD di proteine a struttura incognita, eseguendo un fit secondo l’equazione:
[
q ( l ) ] = c a [ q a ( l ) ] + c b [ q b ( l ) ] + c r[ q r( l ) ]
{kind=link}