CEFALOPODI ASPETTI
GENERALI - MORFOLOGIA e CLASSIFICAZIONE
Generalità
I
Cefalopodi sono la classe più evoluta fra le sei che costituiscono il phylum Mollusca;
attualmente, nella Classe Cephalopoda sono
compresi seppie, polpi e calamari.
Questi
animali, che vivono esclusivamente in ambiente marino, presentano simmetria
bilaterale. Soltanto i Nautiloidea, fra i viventi, possiedono una
conchiglia esterna, che si presenta avvolta e divisa in camere; negli altri
Ordini la conchiglia è interna, calcarea, ridotta ad un sottile filamento
corneo, o è addirittura assente. Il piede è trasformato in un organo
imbutiforme che, espellendo l’acqua contenuta nel mantello, provvede allo
spostamento a ritroso del mollusco, mediante un getto d’acqua.
Contemporaneamente all’acqua, l’animale espelle i prodotti di rifiuto del suo
metabolismo, i gameti ed ossigena le branchie, che sono
in numero di due per lato. L’apparato digerente consiste in una bocca dotata di
un becco corneo e di una radula, a cui seguono un esofago, uno stomaco, un
intestino cieco, un intestino e un ano. Gli occhi sono
particolarmente sviluppati nella maggior parte dei Cefalopodi attuali, tranne nel Nautilus, che presenta un occhio primitivo
di tipo puntiforme privo di cristallino.
Le
dimensioni sono molto diverse: un esemplare di Nautilus non supera i
Il
sistema nervoso presenta una concentrazione di gangli nella regione cefalica.
La separazione tra i sessi è costante. Le uova deposte sono telolecitiche, a
segmentazione discoidale. Il periodo embrionale è lungo e manca lo stadio
larvale. Lo studio degli ottopodi attuali dimostra che il loro sistema nervoso
è ben organizzato, in modo da poter far tesoro dell’esperienza; le femmine, inoltre,
sono in grado di accudire e proteggere la prole.
La
maggior parte delle specie è predatrice carnivora, si nutre di pesci e
crostacei, mentre le forme che vivono in prossimità dei fondali si nutrono di
crostacei, piccoli pesci e molluschi.
I
Cefalopodi attuali si dividono in due classi: Dibranchiati, dotati di un solo
paio di branchie e di otto o dieci tentacoli, di occhi
sviluppati e di sacco dell’inchiostro. I Tetrabranchiati possiedono una
conchiglia esterna, due paia di branchie, molte braccia, occhi assai primitivi
e sono privi del sacco dell’inchiostro. Dal punto di vista paleontologico, la
classe dei Cefalopodi si divide in tre sottoclassi: Nautiloidea, Ammonoidea
e Coleoidea.
I più
antichi rappresentanti dei Cefalopodi costituiscono la sottoclasse Nautiloidea,
che compare nel Cambriano, è diffusa in tutto il Paleozoico con i numerosi
ordini e innumerevoli forme, si riduce fortemente nel Mesozoico, quando subisce
l’esplosione della sottoclasse Ammonoidea, si riduce ulteriormente nel
Cenozoico ed è residuale nel Quaternario. Attualmente
è vivente il solo genere Nautilus, rappresentato da cinque o sei specie
a seconda dei vari autori: N. pompilius, N. macromphalus, N.
stenomphalus, N. scrobiculatus e N. repertus; la conoscenza della loro anatomia è
utilizzata per deduzione per interpretare l’anatomia delle forme fossili.
Il Nautilus
attuale vive nel Pacifico sud occidentale, dalle isole Fiji allo stretto di
Malacca.

![]()

 Phylum Mollusca
Phylum Mollusca
Classi Rostroconchia
Aplacophora
Pelecypoda
Scaphoda
Polyplacophora
Monoplacophora Cephalopoda Gastropoda
Cephalopoda
![]()

Sottoclassi
Bactritoidea Ammonoidea
(Devoniano sup.-Triassico, (Dev. Inf. – Creta) ~ 30 generi)
Nautioloidea
(Cambriano sup.-attuale;

![]()
 ~ 700 generi)
~ 700 generi)
 Coleoidea
Coleoidea
(Dev. Inf.-Att.,~300 g.)
Ordini
Endoceratida Nautilida
Oncoceratida Discosorida
Tarphyceratida Orthoceratida
Schema tassonomico del phylum Mollusca (secondo Pojeta jr, in Boardman, Cheetham & Rowell, 1987)
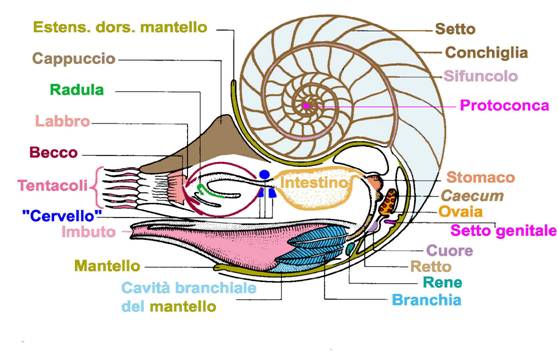
fig. 1 –
Sezione equatoriale della conchiglia di un Nautilus.
La conchiglia è composta da
carbonato di calcio nella forma cristallina d’aragonite, e da sostanze
organiche. È divisa in tre strati: uno esterno
organico, il «black film» degli autori anglosassoni, uno strato porcel-lanaceo
e uno madreperlaceo.
Lo strato porcellanaceo è composto nella sua parte
esterna da microscopici cristalli di aragonite, uniti
tra loro da una matrice di conchiolina scura. Periodi di non deposizione
lasciano un’impronta sullo strato porcellanaceo, formando linee o strie
d’accrescimento.
Lo strato madreperlaceo costituisce i setti ed è
formato da lamelle di cristalli cilindrici d’aragonite alternati a conchiolina.
La conchiglia esterna, sia essa diritta o avvolta,
è riconducibile ad un cono, variabile nelle dimensioni e nella curvatura;
generalmente presenta simmetria bilaterale. Si possono distinguere diversi tipologie di guscio: si parla di gusci ortoconi se
diritti, cirtoconi se curvi, longiconi se lunghi ed affusolati, breviconi se
corti e tozzi. La maggior parte delle conchiglie avvolte mostra un avvolgimento
secondo una planispirale a simmetria bilaterale. Ciascun giro a 360° si dice giro di spira. Il centro di una conchiglia planispirale è
detto ombelico. Le conchiglie con ombelico ampio che mostrano tutti i giri sono
dette evolute, quelle in cui l’ombelico è piccolo e l’ultimo giro ricopre i
precedenti sono dette involute. Esistono vari tipi di sovrapposizione dei giri di spira, e di conseguenza una gradazione tra le forme
evolute e quelle involute.
Alcune conchiglie di Cefalopodi del Siluriano, del
Devoniano e del Mesozoico si avvolgono in modo asimmetrico, e vengono per
questo motivo dette torticone. Un caso estremo di avvolgimento è rappresentato da un genere eteromorfo
dell’ammonite cretacea Turrilites, che esternamente è simile ad un
gasteropode.
Le forme ricurve o avvolte nelle quali il ventre è sul lato convesso esterno sono dette esogastriche, mentre
quelle in cui la parte dorsale è sul lato convesso, sono dette endogastriche. Anche la posizione relativa del seno iponomico aiuta alla
definizione di conchiglie esogastriche ed endogastriche.
Anteriormente, nella parte più ampia della conchiglia, è situata una camera indivisa
che ospita le parti molli, e che è detta camera d’abitazione. Durante
l’accrescimento, l’animale si sposta in avanti e costruisce dietro di sé, nella
parte posteriore della camera d’abitazione, un nuovo setto. La forma più
semplice di camera d’abitazione, detta «aperta», si espande in modo uniforme
dalla base fino all’apertura, e il suo diametro è superiore rispetto a qualunque
altra sezione della conchiglia.
Un’apertura che presenta inizialmente un diametro
maggiore che successivamente diminuisce si definisce
«contratta».
Il caso opposto è quello dell’apertura
«costretta», in cui la camera d’abitazione presenta il diametro minimo poco
prima dell’apertura; a questo restringimento, detto collo, segue immediatamente
una svasatura.
L’ornamentazione esterna è variabile: la più
comune e semplice è rappresentata da linee di accrescimento
con andamento più o meno perpendicolare all’asse longitudinale. Si riscontrano
in alcuni casi le cosiddette striae, sottili solchi paralleli
intervallati da linee rilevate, che possono presentare un andamento parallelo o
normale all’asse longitudinale.
Le coste sono strutture solitamente più
distanziate, sempre rilevate, con andamento in prevalenza longitudinale. Le
annulazioni costituiscono ispessimenti trasversali. Si possono poi trovare ulteriori ornamentazioni: nodi, spine, tubercoli, e molte
altre, specialmente nella sottoclasse Ammonoidea.
L’apertura presenta un’insenatura più o meno profonda, il seno iponomico, che si trova
sul lato ventrale e favorisce la fuoriuscita dell’imbuto. Il margine aperturale
è detto peristoma.
La parte del guscio suddivisa in camere da setti
trasversali è detta fragmocono. Questo prende inizio da una camera
apicale detta protoconca, alla quale si dedica particolare attenzione in
quanto rappresenta lo spazio fra l’apice del fragmocono ed il primo setto, e la
sua forma può essere piuttosto varia. Il paleontologo tedesco Ristedt nel
In molti Nautiloidi l’estremità apicale del
sifuncolo, detta caecum, è saldata al fondo della
camera iniziale e sulla parete esterna di molte forme rimane una piccola
depressione, detta cicatrix.
Le parti interne della conchiglia
Internamente, la conchiglia è divisa in camere da
diaframmi detti setti. Una conchiglia di Nautilus adulto presenta un
numero di camere variabile fra 33 e 36. Lo spazio fra l’ultimo setto e
l’apertura occupa circa 1/3 - ½
dell’ultimo giro. I setti sono composti da aragonite
madreperlacea e sono concavi verso la camera d’abitazione, che si trova in
posizione adorale.
I setti diventano tangenti alla parete della
conchiglia esterna formando una «linea di sutura», che è una linea
formata dall’unione del setto con la parete laterale della conchiglia, molto
importante per la definizione delle selle e dei lobi. Le selle sono formate
dalle ripiegature del setto rivolte verso l’apertura, mentre i lobi sono le
stesse strutture che si allontanano da essa. Nella
sottoclasse Ammonoidea possono essere di vari ordini. La linea di sutura è ben
visibile nei modelli interni e può essere semplice o complessa, presentando
profonde ondulazioni. Queste ripiegature hanno probabilmente la funzione di
irrobustire il fragmocono al fine di contrastare la pressione idrostatica
esterna gravante su di esso.
Le linee di sutura vengono
rappresentate sviluppandole sul piano. Essendo simmetriche, solitamente se ne
proietta soltanto metà, procedendo dalla parte ventrale verso quella dorsale.
Per convenzione, si è soliti marcare la linea mediana di simmetria con una
freccia rivolta verso l’apertura.
Il sifuncolo è un tubicino che collega tutte le
camere attraversando i setti, generalmente in posizione centrale nei nautiloidi,
marginale negli ammonoidi e coleoidi. Nel sifuncolo si possono distinguere un
endosifuncolo costituito da parti molli, comprendenti i vasi sanguigni, ed un
ectosifuncolo, che è costituito dai collaretti settali
e dagli anelli di connessione che collegano i collaretti fra loro. Nel genere Nautilus, la parte costituita dai
tessuti molli è detta corda sifonale, o sifuncolare, mentre la parte esterna,
inorganica e più o meno mineralizzata, è detta
ectosifuncolo. Quest’ultimo generalmente è poroso e composto da
carbonato di calcio e conchiolina, mentre lo spazio ad esso interno si
definisce, per convenzione, endosifuncolare.
Se i collaretti sono ripiegati verso l’apice sono detti retrocoanitici, se sono rivolti verso
l’apertura si dicono procoantici. Possono essere inoltre più
o meno sviluppati ed assumere diverse morfologie. Il sifuncolo di alcuni Cefalopodi fossili può presentare depositi
endosifuncolari. Questi depositi si dicono episettali se si trovano sulla parte
anteriore del setto, iposettali se si trovano sulla parte
posteriore, murali se si trovano sulle pareti laterali delle camere.
Questi depositi camerali hanno probabilmente la funzione di bilanciare la spinta verso l’alto che il fragmocono subisce a causa della
presenza di gas all’interno delle camere, e quindi mantenere un assetto di vita
orizzontale, soprattutto per le forme longicone. Vari autori hanno ipotizzato
l’esistenza di un mantello camerale per spiegare la presenza di questi
depositi, ma l’unica forma attualmente vivente con la
quale si possono fare confronti non presenta strutture di questo tipo. Nelle
forme planispirali il problema della stabilità è in parte
risolto perché il centro di gravità (baricentro) e quello d’equilibrio
si trovano sullo stesso asse.
Gli adattamenti idrostatici che regolano la
posizione dell’animale, in modo tale che ogni movimento sia sempre
perfettamente controllato, sono molto importanti. I sistemi grazie ai quali i
Cefalopodi dotati di conchiglia esterna regolano la loro posizione sono due:
aggiungendo e sottraendo liquido dalle camere del
fragmocono, oppure aggiungendo e sottraendo gas.
Nel primo caso è stato dimostrato che le nuove
camere sono in origine piene di liquido molto simile
per composizione all’acqua marina; in corrispondenza con la calcificazione di
un nuovo setto, il sifuncolo rimuove gli ioni di sale da questo liquido,
rendendolo sempre più iposmotico rispetto al liquido all’interno del sifuncolo.
Le differenze di osmolarità creano un gradiente
osmotico che causa il movimento del liquido dalla camera verso il sifuncolo.
Con l’aumento della profondità, la pressione idrostatica si oppone al movimento
del liquido verso il sifuncolo, perché la pressione all’interno di esso è in equilibrio con la pressione idrostatica. Per questa ragione, il nautiloide, per poter eseguire l’operazione
di svuotamento, migra verso la superficie, dove la pressione idrostatica è più
bassa.
A profondità maggiori, la differenza di pressione
fra la camera e l’interno del sifuncolo è all’incirca di 30 atmosfere. Lo
svuotamento delle camere avverrà solo se la pressione osmotica prodotta dalla
concentrazione salina sarà maggiore della pressione
idrostatica, e il grado di svuotamento dipenderà dalla differenza fra questi
due valori. Se la pressione idrostatica supera la
pressione osmotica, le camere si svuoteranno molto lentamente.
Nel secondo caso si considerano le camere piene di
gas, e quindi si ipotizza che il corpo dell’animale
sia solidale alla conchiglia lungo due linee sinuose sul fianco, ma non sulla
superficie concava dell’ultimo setto; un volume riempito di gas tra questo
setto ed il corpo potrebbe essere alterato dalla contrazione muscolare, in modo
da comprimere o rarefare il gas, aumentando o diminuendo in questo modo la
pressione del gas contenuto nelle camere del fragmocono.
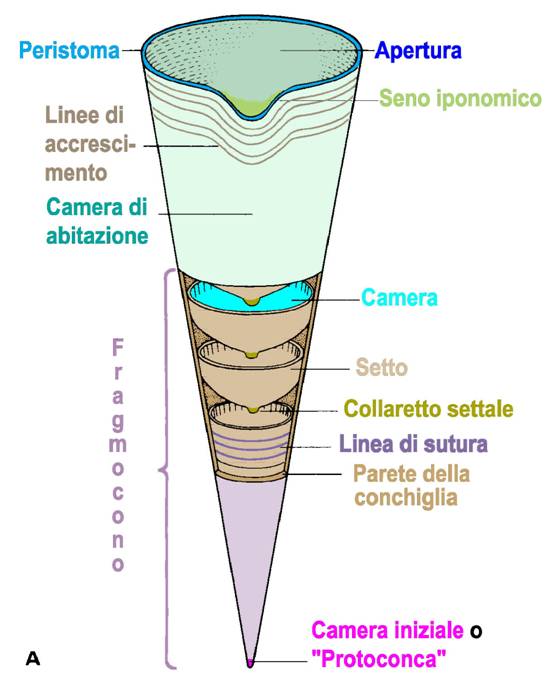
Fig. 2 – Sezione e nomenclatura di un ortocono.
SINTESI GEOLOGICA DELLA SARDEGNA
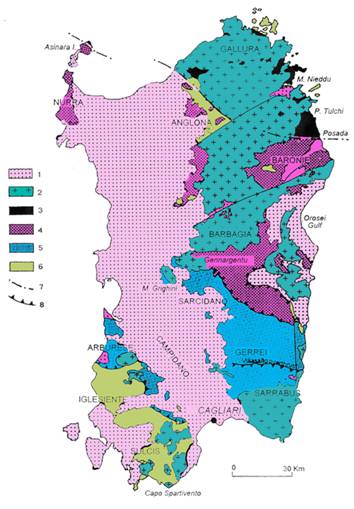
Fig. 1 - schema strutturale del basamento sardo. 1: copertura post-ercinica; 2: batolite ercinico; 3: complesso ad
alto grado di metamorfismo; 4: falde interne; 5: falde esterne; 6: zona
esterna; 7: linea Posada – Asinara; 8: falde principali.
ORDOVICIANO
Le unità litostratigrafiche della
sequenza ordoviciana della Sardegna sud occidentale sono state descritte da
Leone et alii nel 1991, con aggiornamenti da Leone et al., 1998; per
riassumere, queste unità sono costituite da:
FORMAZIONE
DI MONTE ARGENTU («PUDDINGA» auct.)
Nella Sardegna meridionale, la
successione autoctona inizia con conglomerati rossastri, siltiti ed arenarie
(“unità a”, Cocozza e Leone, 1977; “unità a1-a3”, Hamman et al., 1990). Dal
punto di vista litologico, la formazione di Monte Argentu può essere suddivisa
in tre unità (Laske et al.,
1994): Membro di Punta sa Broccia, Membro di Riu is Arrus e Membro di Medau
Murtas.
Il membro di Punta sa Broccia
è composto principalmente da conglomerati rossastri per lo più non fossiliferi,
con strati di siltiti ed arenarie (“unità a”, Laske & Bechstadt, 1987-1989;
“unità a1”, Hamman et al., 1990).
In molte località dell’Iglesiente, e
in particolar modo nell’area di Nebida e Bacu Abis, si notano intercalazioni di
brecce calcaree, in parte dolomizzate, nei conglomerati basali, che si
sovrappongono alla Discordanza Sarda; seguono siltiti e scisti dello spessore
di
Alla base di questo membro, nella
regione del Sulcis e in alcune zone dell’Iglesiente, sono presenti spessi
strati di conglomerato. Inoltre, nell’area di Narcao e Rosas, le siltiti sono
intercalate da calcari, in parte dolomizzati, in sottili strati o “lenti”
(Poll, 1966; Fiori, 1977). Il contenuto fossilifero è scarso sia nella zona
superiore sia nella zona inferiore di questo membro.
Membro di Riu is Arrus
Il membro precedentemente
descritto è sovrastato da siltiti grigie localmente fossilifere di una potenza variabile fra 20 e
Membro
di Medau Murtas
Le siltiti grigie e gli scisti precedentemente menzionati sono seguiti da una sequenza di
oltre
Ad eccezione di rare tracce fossili
nell’area di Bacu Abis (Pillola, 1981; Loi et al., 1995) il contenuto fossilifero
della Formazione del Monte Argentu è scarso; solo nel Membro di Riu is Arrus
Hamman et al., 1990 riportano un insolito trilobitomorfo, Tarricoia
arrusensis.
A causa della mancanza di fossili
biostratigraficamente significativi, la sequenza di
Monte Argentu non può essere datata direttamente. Leone et al. (1988) riportano
l’inizio della sedimentazione di questa formazione nel primo Caradociano
(Souldeyano).
FORMAZIONE DEL MONTE ORRI
Questa formazione consiste
quasi esclusivamente di una sequenza silico-clastica di siltiti grigio-verdi.
Nella sezione tipo mostra uno spessore di circa
Nell’area di Fluminimaggiore la
formazione di monte Orri contiene fossili marini tra i quali ricordiamo
brachiopodi (articolati e inarticolati), bivalvi e trilobiti.
L’ambiente deposizionale è
caratterizzato da una sedimentazione molto irregolare, in prevalenza di
materiale sabbioso con ichnofossili tipo “Skolithos”. Il livello del mare ha
subito un rapido abbassamento, inoltre la concentrazione di grandi quantità di
fauna marina solo in alcuni livelli e l’abbondanza di lingulidi in altri,
potrebbe far pensare ad una variazione di salinità (Havliček et al., 1987).
formazione di portixeddu
La formazione consiste principalmente
di siltiti e scisti grigio-scuri altamente
fossiliferi, contenenti localmente noduli fosfatici e di pirite. Lo spessore
stimato di questa litologia varia dai 50 ai
formazione di domusnovas
Questa formazione comprende due unità
litologiche: il membro di Maciurru, per lo più
arenaceo e scarsamente fossilifero, e il membro di Punta S’Argiola, marnoso.
Una caratteristica peculiare di quest’ultimo è il colore rosso delle rocce, che
permette l’identificazione nonostante lo spessore del
primo membro sia estremamente ridotto o assente. La colorazione rossa delle
argilliti, derivante dalla diagenesi dell’ematite, indica un alto contenuto di componenti ricchi in ferro (chamosite-gothite). Il primo
membro è scarsamente fossilifero; nella parte inferiore della sezione si
trovano brachiopodi, briozoi e crinoidi. Cocozza e Leone (1977) suggeriscono
una sedimentazione di tipo regressivo per il membro di Punta S’Argiola, in relazione alla glaciazione dell’Ordoviciano superiore;
molte evidenze sedimentologiche suggeriscono infatti condizioni di alta energia
a tendenza regressiva. La parte mediana e quella inferiore
del secondo membro è sicuramente la più ricca in fossili: briozoi, crinoidi,
cistoidi, trilobiti, brachiopodi. Nella parte superiore del membro si trovano
numerosi resti di cefalopodi ortoconi, dei quali è conservato il sifuncolo con
ampi depositi endosifuncolari, tipici degli Actinoceratidi ed Endoceratidi.
FORMAZIONE DI RIO SAN MARCO
Questa formazione è costituita da
depositi clastici, per lo più da torbiditi, ed è suddivisa in quattro membri:
Punta Arenas, Cuccurruneddu, Serra Corroga e Girisi. Il membro di Punta Arenas
è assai sottile, con strati brecciati e conglomerati di colore verde, in cui
prevalgono clasti di origine vulcanica (Beccaluva et
al., 1981), alternati con siltiti grigio verdi e scisti silicei. Negli
scisti neri si possono trovare rari frammenti di briozoi, mentre, negli strati
presentanti una breccia molto fine, si rinvengono due specie di graptoliti, per
il cui riconoscimento occorre fare riferimento, rispettivamente, a Dr. H.
Jaeger e Dr. P. Storch. La presenza di materiali vulcanici in quest’unità
suggerisce una contemporanea attività vulcanica nella zona, anche se la
sorgente di questi clasti rimane comunque molto
difficile da stabilire. La varietà della facies è probabilmente in relazione
con un’attività vulcanica abbastanza complessa, sia sottomarina sia subaerea. Inoltre, si possono descrivere fenomeni
d’eruzioni magmatiche all’interno di sedimenti plastici. Caratteristici di
questo membro sono anche alcuni livelli di origine
torbiditica. I responsabili di questi processi potrebbero
essere stati i movimenti tettonici, come suggerito dall’incremento della
batimetria, ma anche processi erosivi, influenzati dalla regressione
glacioeustatica ordoviciana (Berry & Boucot, 1973; Brenchley & Newall,
1980; Brenchley, 1988). Il membro di Cuccurruneddu ha una litologia che
consiste in una ripetizione ritmica di strati di torbiditi pelitico-arenacee,
di un colore che varia dal grigio al grigio chiaro. Gli intervalli arenacei
sono composti esclusivamente da quarziti micacee. L’unità è praticamente
priva di macrofossili, mentre si possono notare molti tipi di ichnofossili. In
alcuni orizzonti pelitici sono conservati acritarchi e chitinozoi (Del Rio et
al., 1979).
Anche in questo membro, come nel precedente, la
deposizione è di tipo torbiditico. Mancano tuttavia studi dettagliati ed
analisi sedimentologiche approfondite, ed è quindi
impossibile mettere in relazione queste torbiditi con una facies ben definita.
Il membro di Serra Corroga presenta una litologia di siltiti
molto fini di colore grigio scuro e verde scuro, con laminazioni molto
sottili di tipo varvale. Queste strutture sono certamente correlate a correnti
di torbida di bassa densità. La porzione inferiore del membro di Girisi
presenta un’alternanza di siltiti grigio scure ed
argilliti con arenarie molto fini, mentre la parte superiore è formata da
siltiti e argilliti altamente micacee e per lo più massive, interstratificate
con arenarie molto fini. I fossili sono molto rari: si trovano soltanto
frammenti di modelli esterni di brachiopodi articolati e di crinoidi, immersi
in una matrice pelitica scura. Nella parte superiore sono stati rinvenuti anche
acritarchi (Del Rio, Leone & Pittau, 1979). I sedimenti di questa
formazione hanno molte caratteristiche in comune con le facies glaciomarine
della parte superiore della successione ordoviciana europea, e sono interpretati
come materiali eterogenei, trasportati attraverso ghiacci galleggianti e
depositatisi in ambiente di mare aperto all’interno della normale
sedimentazione pelagica (Brenchley & Štorch, 1989).
SILURIANO E
DEVONIANO
Per quanto riguarda gli
affioramenti siluro-devoniani della Sardegna sud occidentale, sono state proposte tre unità litostratigrafiche formali:
Formazione di Genna Muxerru; Formazione di Fluminimaggiore; Formazione di Mason
Porcus. La successione di questi terreni è costituita da una sequenza sedimentaria
di circa
La formazione di Fluminimaggiore
presenta un’alternanza di livelli calcarei neri d’aspetto lenticolare,
presentanti solitamente numerosi nautiloidi, intersecati da peliti scure e
rocce argillose. Prevalentemente, questi corpi lentiformi sembrano non aver
sofferto alcun clivaggio, e la maggior parte dei fossili contenuti ha
conservato un aspetto tridimensionale (Gnoli et al., 1979). Lo spessore esatto della
formazione , a causa di intensi fenomeni tettonici, è
difficilmente misurabile, anche se è possibile stimarne una potenza di circa
La formazione di Mason Porcus è
composta per la maggior parte di carbonati, e costituisce la più importante
espressione topografica delle formazioni siluro-devoniane dell’area
Iglesiente-Sulcis. La totalità delle formazione non è
esposta in una singola sezione dell’area tipo, e per coprire l’intera
formazione si è dovuto fare riferimento a diverse sezioni; la litologia
principale è quella dei calcari nodulari, spesso finemente stratificati ed
anche massivi, alternantisi con siltiti e scisti ad emitrochi e frammenti di
steli di crinoidi. All’interno della sequenza è possibile osservare un “Mound”
carbonatico a Stromatactis di aspetto lenticolare, affiorante a Monte Padenteddu nel
Sulcis meridionale. Localmente questa formazione può raggiungere i
Le rocce fossilifere siluro-devoniane
sono abbastanza diffuse nella Sardegna sud-orientale
ed appartengono all’Unità del Sarrabus e all’Unità del Gerrei, le unità che
hanno subito in misura minore il metamorfismo della cintura a falde della
Sardegna. La successione paleozoica della regione del Gerrei è costituita da
una potente formazione silico-clastica (Arenarie di San Vito) contenente
acritarchi Cambro-Tremadociani, che sono sovrastati (“Discordanza Sarrabese”)
da metavulcaniti acide (“Porfiroidi”), d’età Pre-Caradociana. I sedimenti
fossiliferi del Caradoc-Ashgill, consistenti di conglomerati, arenarie e siltiti, giacciono su metavulcaniti
più vecchie. Nella parte alta della sequenza si trovano calcari totalmente o parzialmente
silicizzati. La successione prosegue prima con argilliti liditiche nere
(LGS = Lyditic black shales) e calcari di età
siluriana (Ockerkalk facies) ed ancora con calcari marnosi contenenti
dacrioconaridi (Devoniano medio-inf.). La sequenza Devoniano sup.- Carbonifero
inf. termina con calcari pelagici (Calcari a Clymeniae Auct.), conglomerati e
siltiti legate a un complesso flyscioide
dell’Orogenesi Ercinica.
La sezione studiata è esposta sul
versante meridionale della collina di quota
 Fig.
2 - schizzo geologico dei dintorni del paese di San
Basilio. La sezione studiata è indicata dalle lettere A-B e B-C.
Fig.
2 - schizzo geologico dei dintorni del paese di San
Basilio. La sezione studiata è indicata dalle lettere A-B e B-C.
La sequenza delle litologie, dal
basso all’alto, è la seguente (fig. 3):
-
Siltiti
fossilifere grigio-verdi dell’Ordoviciano superiore;
-
argilliti nere (cataclastiti) con liditi (Siluriano inferiore?) (
-
circa
-
argilliti grigio verdi (cataclastiti) (circa
-
“Ockerkalk”
(
-
sottili cinture di cataclastiti;
-
spessa sequenza (
-
argille grigio verdi (
-
circa
-
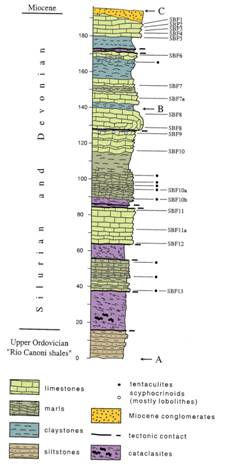 contatto tettonico seguito da un’anticlinale
inversa composta da calcari a facies Ockerkalk, seguite da argilliti grigio
verdi, che passano gradatamente verso l’alto a marne e calcari nodulari (
contatto tettonico seguito da un’anticlinale
inversa composta da calcari a facies Ockerkalk, seguite da argilliti grigio
verdi, che passano gradatamente verso l’alto a marne e calcari nodulari (
-
sottile cintura a cataclastiti
-
argille grigio verdi (
-
calcari argillosi blu verdi (
Fig. 3 – colonna stratigrafica della sezione di San Basilio Fenugu.
La sequenza paleozoica termina verso
la cima della collina ed è coperta da sedimenti clastici miocenici.
Biostratigrafia
L’età della fauna a conodonti è
compresa tra il tardo Siluriano ed il Frasniano, ma alcuni intervalli non sono
documentati nella sezione di San Basilio Fenugu. Sono state distinte le
seguenti biozone:
Zona ad Ancoradella ploeckensis (Ludlow,
Siluriano superiore)
La zona ad A. ploeckensis è
stata riscontrata nel campione SBF 5 per la presenza del marker Ancoradella
ploeckensis Walliser, 1964. La fauna comprende anche Ozarkodina exc. excavata (Branson & Mehl, 1934), Kockelella v.
variabilis Walliser, 1957 ed Oulodus siluricus (Branson & Mehl,
1934).
Zona a Polygnathoides siluricus
(Ludlow, Siluriano superiore)
È stata riscontrata nei campioni SBF
3 e SBF 4. Oltre al marker, Polygnathoides
siluricus Branson & Mehl, 1934, la fauna comprende Oz. exc. excavata (Branson & Mehl, 1934), Oz. confluens (Branson & Mehl, 1934) e Coryssognathus
dubius (Rhodes, 1953).
Zona a Ozarkodina
crispa (Ludlow, Siluriano superiore)
È stata riscontrata nel campione SBF
2. L’associazione è abbastanza ricca e comprende Ozarkodina crispa
(Walliser, 1964), Oz. rem. eostenhornensis s.l., Oz. snajdri
(Walliser, 1964), Oz. exc. excavata (Branson
& mehl, 1934), Oulodus el. elegans
(Walliser, 1964), Pelekysgnathus cf. index
(Klapper & Murphy, 1975), Pseudooneotodus beckmanni (Bischoff
& Sannemann, 1957), Ps. bicornis Drygant,
1984, Belodella resima Philip, 1965 e Panderodus recurvatus (s. l.).
Intervallo ad Ozarkodina
remscheidensis (Přídolí, Siluriano superiore)
È la biozona più diffusa nella
sezione, essendo stata riscontrata negli intervalli SBF 1, 7A,
8, 9, 11, 11A e 12. Inoltre, è possibile affermare che i campioni SBF 8, 11 e
12 appartengono alla parte superiore della zona per la presenza di Oz. rem. Remscheidensis (Ziegler, 1960). La fauna
include anche Oz. rem. eosteinhornensis (Walliser,
1964), nei morfotipi a e b, Oz. exc. excavata
(Branson & Mehl, 1934), Oz. confluens (Branson
& Mehl, 1934), Coryssognathus dubius (Rhodes, 1953), Oul. el. elegans (Walliser, 1964), Oul. siluricus
(Branson & Mehl, 1934), Pseudooneotodus beckmanni (Bischoff
& Sannemann, 1957) e Ps. bicornis Drygant,
1984.
Zona a Oulodus
elegans detortus (Pridoli, Siluriano superiore)
Viene riscontrata nei campioni SBF 7, SBF
9 e SBF 10. Oltre al marker, la fauna include Oz.
rem. remscheidensis (Ziegler, 1960), Oz. rem. eosteinhornensis (Walliser, 1964), Oz. exc. excavata (Branson & Mehl, 1934), Coryssognathus
dubius (Rhodes, 1953), Ps. bicornis
Drygnant, 1984 e Belodella resima (Philip, 1963).
Zona delta (Lochkoviano,
Devoniano inferiore)
È rappresentata dal campione SFB 6.
La fauna include Ancyrodelloides limbarcarinatus Murphy & Matti,
Zona a pesavis (Lochkoviano,
Devoniano inferiore)
Il campione SBF 10b
presenta un contenuto faunistico molto povero; tuttavia, la presenza di Oz.
pandora (Murphy & matti, 1982) morfologia e, suggerisce una datazione alla zona pesavis.
È anche presente Belodella sp.
Intervallo a kitabicus-excavatus (Emsiano,
Devoniano inferiore)
Il campione SBF 10a presenta un
contenuto fossilifero molto scarso e mal conservato. Il
ritrovamento di un paio di frammenti di Polygnathus cf. dehiscens e qualche esemplare di Pandorinellina
steinhornensis permettono di attribuirlo all’intervallo a kitabicus-excavatus.
La zona superiore a rhenana (Frasniano,
Devoniano sup.)
La zona è identificata dal campione
SBF 13 per la presenza di Icriodus alternatus
alternatus Branson & Mehl, 1934 e di Ancyrodella ioides Ziegler,
1958. L’associazione include anche Ancyrodella cf. buckeyensis
Stauffer, 1938, Ancyrognathus triangularis Youngquist, 1945, Palmatolepis
hassi Müller & Müller, 1957 e Polygnathus procerus, Sannemann,
1955.
Conclusioni
Sono state documentate cinque biozone
del Siluriano superiore (A. ploeckensis, Pol. siluricus, Oz. crispa, Oz. remscheidensis,
Oul. el. detortus), tre del Devoniano inferiore
(delta, pesavis, kitabicus-excavatus) ed una del Devoniano
superiore (rhenana).
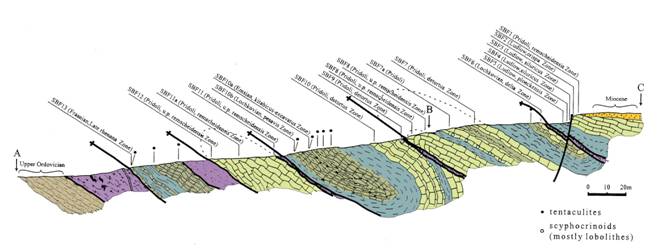 Di seguito la sezione geologica A-B-C riportata nello schizzo geologico
di SBF
Di seguito la sezione geologica A-B-C riportata nello schizzo geologico
di SBF
PARTE SISTEMATICA
La fauna descritta è stata raccolta
dal dott. C. Corradini nel 1991 nella località San Basilio Fenugu, ed
appartiene alla Sezione di Cuccurru Fenugu (cartografia regionale in scala 1:
10.000, Tavola S. Basilio, Foglio 225 ISO). Gli esemplari esaminati provengono
da meta-calcareniti fossilifere corrispondente al livello 12 della sezione di
fig. 3, riportata nella parte introduttiva. La fauna consiste di 14 cefalopodi
appartenenti ai generi Temperoceras, Michelinoceras, Kopaninoceras,
Columenoceras ed Oonoceras ed è stata datata al Ludlow superiore
tramite biozone a Conodonti: crassa -
latialata (Ludlow sup.).
Genere Michelinoceras Foerste, 1932
Specie tipo - Orthoceras michelini Barrande, 1866 per
designazione originale.
1932
Michelinoceras Foerste, p. 72.
1944
Michelinoceras Foerste - Shimer
& Shrock, p. 537.
1945
Michelinoceras Foerste - Flower,
p. 682.
1949
Michelinoceras Foerste - Miller
& Youngquist, p. 28.
1952 Michelinoceras Foerste
- Basse, p. 492.
1956 Michelinoceras Foerste
- Schmidt, p. 50.
1960 Michelinoceras
Foerste - Müller, p. 103.
1962 Michelinoceras
Foerste - Flower, pp. 10/1.
1962 Michelinoceras
Foerste - Balashov & Zhuravleva,
p. 83.
1964 Michelinoceras
Foerste - Sweet, pp. 225/6.
1965 Michelinoceras
Foerste - Gordon, pp. 105/6.
1966 Michelinoceras
Foerste - Babin, p. 321.
1968 Michelinoceras
Foerste - Ristedt, pp. 243-244.
1968 Michelinoceras
Foerste - Shimansky, pp. 52/3.
1971 Michelinoceras
Foerste - Kiselev, pp. 43/4.
1972 Michelinoceras
Foerste - Barskov, pp. 35/6.
1974 Michelinoceras
Foerste - Balashov & Zhuravleva,
p. 107.
1977 Michelinoceras
Foerste - Xu, p. 541.
1977 Michelinoceras
Foerste - Serpagli & Gnoli,
p. 161.
1978 Michelinoceras
Foerste - Zhuravleva, pp. 49-50.
1984 Michelinoceras
Foerste - Dzik, p. 125.
1987 Michelinoceras
Foerste - Chen, pp. 141/2.
1992 Michelinoceras
Foerste - Kiselev & Gnoli, p.
74.
1994 Michelinoceras
Foerste - Müller, p. 160.
1998 Michelinoceras
Foerste - Histon, p. 28.
Osservazioni - Michelinoceras è uno dei generi a più ampia
diffusione nel Siluriano. È riportato dalla letteratura in: [Parte settentrionale
del Gondwana] ovvero nelle attuali Repubblica Ceca
(Barrandium), Francia (Bretagna occidentale, Provenza), Germania, Sardegna
(Italia), Caucaso (Gnoli, 2002), Nord America, Tibet (Cina), Cina, Giappone e
Australia.
Michelinoceras cf. currens (Barrande, 1866)
(Tav. 1, fig.1)
1857 Orthoceras simplex Desnoy - Meneghini,
tav. C, fig. 4Ab’, 4Ab’’.
1857 Orthoceras submoniliforme Meneghini,
tav. C, fig. 9a (non figg. 9b,
9b’).
1860 Orthoceras currens Barrande,
p. 624 (nomen nudum).
1866 Orthoceras currens Barrande, tav. 221, fig. 26, tav. 222, figg. 15,
16.
1870 Orthoceras currens Barrande, tav. 407, figg. 20-33, tav. 411, figg.
16-18.
1874 Orthoceras currens Barrande, p. 628.
1962 Michelinoceras currens (Barrande)
- Flower, p. 10.
1972 Michelinoceras currens
(Barrande) - Barskov, p. 36, tav. 1, fig. 5,
tav. 2, fig. 5.
Descrizione - Conchiglia ortocona a sezione trasversale
circolare. Solitamente ha un angolo d’espansione di 5 gradi, che a volte può raggiungere i 7. Non è stata osservata alcun’ornamentazione.
La sutura è diritta, semplice ed obliqua. L’altezza delle camere può variare
fortemente: da 4/5 a 2/5 del diametro della conchiglia. I setti sono concavi e
la loro profondità varia da ½ a 1/3 del diametro. Il
sifuncolo è subentrale. Il diametro del foramen settale è circa 1/15 di quello della conchiglia. I
collaretti settali sono ortocoanitici, e la loro
lunghezza è leggermente superiore al diametro del foramen settale. Gli
anelli di connessione sono per lo più cilindrici e a volte si espandono
leggermente nelle camere. Non sono stati osservati depositi.
Osservazioni - Come sottolineato da
Barrande (1874, p. 628) M. currens mostra una grande variabilità nella
lunghezza delle camere, anche nello stesso esemplare. La specie può essere
distinta da M. grande (Meneghini) soprattutto da un più ampio angolo
d’espansione, camere più corte e dal sifuncolo più stretto.
Distribuzione –
Siluriano Superiore della Boemia (Barrande), Asia Centrale, Caucaso e Podolia
(Barskov, 1972), Sardegna sudoccidentale (Gnoli &
Serpagli, 1977) e sud orientale (presente tesi).
Collocazione -
collezione paleontologica del Dipartimento di Scienze della Terra
dell’Università di Modena e Reggio Emilia (cat. n.
24166 – 24167)
Materiale studiato - due esemplari.
Michelinoceras cf. michelini (Barrande,
1866)
(Tav. 1, fig. 2)
1866 Orthoceras michelini
Barrande, tav. 221, figg. 16-17.
1870 Orthoceras
michelini Barrande, tav. 381, figg. 3-14, (secondo Kiselev, 1971non figg.
15-16, tav. 442, figg. 20-23, tav. 447, figg. 12-13).
1874 Orthoceras michelini.
Barrande, p. 642.
1888 Orthoceras michelini.
Barrande - Foord, p. 27.
1918 Orthoceras cf. michelini.
Barrande - Janishevsky,
p. 55, tav. 2, fig.
11.
1924 Orthoceras michelini
Barrande - Zittel-Broili, p. 516,
fig. 1077.
1926 Orthoceras Michelini
Barrande - Foerste, p. 6.
1929 Orthoceras michelini. Foerste, p. 160.
1930 Orthoceras michelini
Barrande - Yü, p. 53.
1932 Michelinoceras michelini
(Barrande) - Foerste, p. 73.
1962 Michelinoceras michelini
(Barrande 1866) - Balashov, p.
83, tav.
10, fig. 15.
1966 Michelinoceras michelini
(Barrande 1866) - Barskov, p. 18.
1968 Michelinoceras michelini
(Barrande 1866) - Kiselev, p. 18.
1968 Michelinoceras
michelini (Barrande 1866) - Ristedt,
pp. 245/6, tav.
1, figg. 1-2, text-fig. 3-1b.
1971 Michelinoceras michelini
(Barrande 1870) - Kiselev, pp.
44/5, tav.
1, figg. 1-5.
1972 Michelinoceras michelini
(Barrande) - Barskov, p. 36, tav.
1, figg.
1, 2, tav. 2, figg. 1-3.
1977 Michelinoceras grande
(Meneghini, 1857) - Serpagli & Gnoli,
pp.
162-5, tav. 2, figg. 1-4, text-fig. 7a
[non tav. 2, figg. 5a, b =
lectotipo
di Columenoceras grande (Meneghini)]
1984 Michelinoceras michelini
(Barrande, 1866) - Dzik, p. 97.
1992 Michelinoceras cf. michelini (Barrande) - Gnoli, p. 266 .
1994 Michelinoceras
michelini (Barrande) - Müller,
p. 161, text-fig.
175a.
1997 Michelinoceras michelini
(Barrande) - Marek in Turek, Marek &
Benes, p. 234.
1999 Michelinoceras michelini
(Barrande) - Histon, & Gnoli,
p. 384, tab.
1.
Neotipo - Campione
illustrato da Barrande (1870) nella tavola 338, figg. 3-16.
Descrizione
- Conchiglia ortocona a sezione circolare. L’angolo d’espansione è di norma molto stretto, solitamente inferiore a 3 gradi. La
camera d’abitazione è leggermente conica oralmente e si sviluppa in lunghezza
per circa 16 volte il suo diametro di base. Non è stata osservata
alcun’ornamentazione. Le suture sono diritte, semplici e trasversali, e variano
da ½ a 1,5 volte il diametro della conchiglia. Si osservano notevoli variazioni
all’interno dello stesso esemplare. I setti sono concavi, e la loro profondità
varia da
Osservazioni - Barrande
raffigurò questa specie (1866, 1870) e la descrisse (1874) attribuendole il
nome Orthoceras michelini, e basò la sua diagnosi soltanto su fram-menti
di fragmoconi adulti. Nel 1932 Foerste istituì il
nuovo genere Michelinoceras, scegliendone come specie-tipo Orthoceras
michelini Barrande. Nel 1968 Ristedt figurò come Michelinoceras
michelini un esemplare giovane in cui erano
conservate la protoconca e le successive 8 camere. Lo stesso Ristedt figurò
(tav. 1, fig. 2) un secondo piccolo campione con 7 camere, appartenenti
probabilmente all’inizio della parte adulta. Infine, Barskov descrisse nel 1972
quattro frammenti di fragmoconi adulti provenienti dal Siluriano
dell’Afganistan e dell’Unione Sovietica.
Distribuzione - Siluriano Superiore della Sardegna (Meneghini,
1857), Boemia (Barrande, 1866, 1870; Ristedt, 1968), Afganistan e Kazakistan meridionale
(Barskov, 1972).
Collocazione -
collezione paleontologica del Dipartimento di Scienze della Terra
dell’Università di Modena e Reggio Emilia (cat. n.
24164 – 24165)
Materiale studiato - cinque esemplari.
Genere Kopaninoceras Kiselev, 1969
Specie
tipo - Orthoceras jucundum Barrande, 1870.
Kopaninoceras cf. thyrsus (Barrande, 1870)
(Tav. 1, fig. 3)
1870 Orthoceras thyrsus Barrande,
tav. 405, figg.
15-18.
1874 Orthoceras thyrsus
Barrande, pp. 555, 556.
1930
Orthoceras thyrsus ?
Barrande - Yü, pp. 54/5, pl. 7,
figg.3a, b.
1962 Michelinoceras thyrsus (Barrande) - Flower, p. 10.
1968 Michelinoceras n. sp. A, Ristedt,
pp. 246, 247, tav. 1, figg. 4a (3?,
4?).
1972 Michelinoceras
thyrsus (Barrande) - Barskov,
p. 35.
1977 Kopaninoceras? thyrsus (Barrande, 1870) - Serpagli
& Gnoli, p.
161, tav. 1, figg. 4a, b.
1991 Kopaninoceras? thyrsus
(Barrande) - Gnoli & Serpagli,
pp. 190,
194,
tav. 3, figg. 4, 5.
1992 Kopaninoceras? cf. thyrsus
(Barrande) - Gnoli, p. 266.
1999 Kopaninoceras thyrsus
(Barrande) - Histon, & Gnoli,
p. 384,
tabl.1.
Descrizione -
Conchiglia ortocona a sezione circolare. Ha un angolo d’espansione molto
stretto, con un’ampiezza inferiore a 4 gradi. La camera d’abitazione era
probabilmente molto lunga. Mostra ornamentazioni formate da linee
d'accrescimento estremamente fini, una o due per
millimetro inclinate di circa 15 gradi in senso dorso ventrale. La sutura è
diritta, semplice ed inclinata come l’ornamentazione. Le camere sono lunghe
metà del loro diametro. I setti sono concavi e la loro profondità misura circa 1/3 del loro diametro. Il sifuncolo è
leggermente più eccentrico ventralmente del suo diametro. Il diametro
del foramen settale misura circa 1/10 di quello
della conchiglia. I collaretti settali sono
leggermente conici, si assottigliano apicalmente ed hanno una lunghezza pari a
1,5 volte il diametro del foramen
settale. Gli anelli di connessione sono cilindrici. Nella parte apicale
d’alcuni esemplari adulti si osservano sottili depositi episettali.
Osservazioni - Restano
alcuni dubbi sulla posizione generica di questa specie. Alcuni autori (Kiselev,
1972, p. 472) sostengono che sia da considerare appartenente al genere Michelinoceras,
ma i nostri esemplari, i cui caratteri concordano con quelli del tipo di
Barrande, hanno collaretti più simili a quelli del genere Kopaninoceras.
Occorre rilevare che Barrande, nella sua descrizione originale d’O. thyrsus (p.
556), si riferisce alle sottili striature affermando che hanno un’inclinazione
di 35 gradi. È tuttavia abbastanza evidente, osservando la tavola 405, fig. 15,
che queste striature sono inclinate di 15-16 gradi.
Distribuzione - Siluriano Superiore della Boemia (Barrande, 1870;
Ristedt, 1968), Siluriano della Sardegna sudoccidentale (Gnoli & Serpagli, 1977) e
sud-orientale (presente tesi).
Collocazione -
collezione paleontologica del Dipartimento di Scienze della Terra
dell’Università di Modena e Reggio Emilia (cat. n.
24168 – 24169)
Materiale studiato - due esemplari.
Famiglia Geisonoceratidae Zhuravleva, 1959
Genere Columenoceras
Barskov, 1960
Specie tipo - Orthoceras columen Barrande, 1868.
Osservazioni - Il genere Columenoceras era stato istituito
originariamente da Barskov (1960) con lo specie tipo Orthoceras columen
Barrande, ed era stato attribuito alla famiglia Pseudorthoceratidae. Il
genere fu poi preso in considerazione da Sweet (1964, p. K237) come
rappresentante della famiglia Geisonoceratidae, probabilmente sulla base di caratteri sifuncolari e per la forma dei
depositi endosifuncolari. Infine Dzik (1984, pp. 126, 127) propose una nuova
classificazione basata su un approccio filogenetico, secondo la quale il genere
Columenoceras apparterrebbe alla famiglia Orthoceratidae.
Pensiamo,
seguendo la linea della classificazione del «Treatise» (Sweet, 1964), che i
caratteri che permettono di distinguere questo genere dagli altri della
famiglia Geisonoceratidae siano le dimensioni della conchiglia da medie
a grandi, l’angolo d’espansione molto stretto (2° o meno), un fragmocono
formato da camere tanto lunghe quanto larghe, un ampio sifuncolo ortocoanitico
e la presenza di depositi endosifuncolari “annulari”. L’ornamentazione esterna
della specie tipo è talmente comune fra i cefalopodi ortoconi che non può rappresentare un carattere diagnostico a livello di
genere.
Columenoceras cf. grande (Meneghini,
1857)
(Tav. 2, fig. 1)
1857
Orthoceras grande Meneghini,
pp. 189-191, pl. C, figg. 4A, a,
a’.
1870
Orthoceras grande Meneghini - Barrande,
p. 30.
1949
Orthoceras grande Meneghini - Comaschi
Caria, pp. 203,
335.
non 1977 Michelinoceras
grande (Meneghini) - Serpagli &
Gnoli, pp.
162-165, pl. 2,
figg. 1-4, fig. 7a
(=Michelinoceras michelini (Barrande)).
1977
Columenoceras sp. cf. C. columen (Barrande) - Serpagli &
Gnoli, pp. 186, 187, pl. 3,
figg. 6a-b.
non 1983 Michelinoceras
grande (Meneghini) - Gnoli,
p. 82, pl. 2,
figg.
2, 3 (= Michelinoceras michelini (Barrande)).
1984
Michelinoceras grande (Meneghini) - Dzik,
p. 97.
Descrizione – Conchiglia ortocona molto sottile di grandi dimensioni,
con un diametro fino a
Distribuzione – la specie sembra essere limitata stratigraficamente al
Wenlock superiore della Sardegna sud occidentale e al Siluriano della Sardegna
sud orientale.
Collocazione -
Collezione Paleontologica del Dipartimento di Scienze della Terra
dell'Università di Modena e Reggio Emilia, cat. n.
24170 - 24171.
Materiale studiato - due esemplari.
Genere Temperoceras
Barskov, 1960
Specie tipo - Orthoceras
temperans Barrande, 1874.
Temperoceras cf. temperans (Barrande)
(Tav. 2, fig. 2)
1866 Orthoceras
temperans Barrande, pl. 226,
figg.
1-4; pl. 230, figg. 7-
9.
1868 Orthoceras
temperans Barrande, pl. 327,
figg.
1-2; pl. 382, figg. 1-
5.
1870 Orthoceras
temperans Barrande, pl. 382,
figg.
1-5; pl. 451, figg. 1-
3.
1874 Orthoceras
temperans Barrande, pp.
658-659.
1960 Temperoceras
temperans (Barrande) - Barskov,
p. 153.
1966 Temperoceras temperans (Barrande)
- Babin, pp. 324-325, pl. 13,
fig. 15; pl. 14, figg. 1a-c.
1972 Temperoceras
temperans (Barrande) - Barskov,
p. 49, pl. 5, figg. 1,
2.
1992 Temperoceras
cf. temperans (Barrande) - Gnoli, p. 266
1999 Temperoceras
temperans (Barrande) - Histon &
Gnoli, p. 384, tab. 1.
Descrizione -
Conchiglia ortocona liscia di grandi dimensioni a sezione trasversale circolare
che si allarga gradualmente secondo un angolo di circa 6°. Il sifuncolo è
largo, centrale, subortocoanitico e leggermente espanso all’interno delle
camere. Il suo diametro massimo è circa 1/10 il diametro della conchiglia e
diminuisce leggermente in corrispondenza dei foramina settali.
I collaretti settali sono subortocoanitici,
leggermente conici, si assottigliano nella parte terminale; la loro lunghezza è
circa 1/3 quella del foramen settale. Le camere sono relativamente brevi, 1/7 il loro diametro. La profondità settale è 1/3 il loro diametro. Lo spessore della conchiglia
è di circa
Osservazioni -
Questa grossa specie è stata accuratamente descritta ed illustrata da Barrande
(1866) e in seguito emendata da Barskov (1960), che l’ha elevata a nuova specie
tipo per il suo nuovo proposto genere Temperoceras.
La diagnosi originale è stata completata in un secondo tempo da Babin (1966),
che ha descritto la presenza di “ostruzioni” sifuncolari.
Distribuzione -
Siluriano superiore della Boemia (Barrande, 1866), di Saint Sauveur-le-Vicomte,
Bretagna nord occidentale (Francia) (Babin, 1966), del Fergana meridionale
(Barskov, 1972), Ludlow medio della Sardegna sud
occidentale e sud orientale (presente tesi).
Collocazione -
collezione paleontologica del Dipartimento di Scienze della Terra di Modena e
Reggio Emilia, cat. n. 24162 - 24163.
Materiale studiato - due
esemplari.
Ordine Oncocerida Flower in Flower &
Kummel, 1950
Famiglia Oncoceratidae Hyatt, 1884,
Genere Oonoceras
Hyatt, 1884
Specie tipo - Cyrtoceras
acinaces Barrande, 1866 per
designazione
successiva, Bassler, 1915.
Oonoceras aff. potens (Barrande,
1866)
(Tav. 3, figg. 1, 2)
1866 Cyrtoceras
potens Barrande, tav. 131,
figg. 10-13.
1867 Cyrtoceras
potens Barrande, pp. 612,
613.
1992 Oonoceras potens (Barrande) - Gnoli, pp. 267/8, figg. 2a, b.
Descrizione - Conchiglia esogastrica ortocona leggermente
compressa di sezione trasversale ovale. Il fragmocono ha setti molto vicini. Le
camere sono corte e misurano circa 1/3 del loro
diametro dorso ventrale, che corrisponde a ½ della lunghezza della conchiglia.
La curvatura delle camere è circa 1/6 del loro
diametro. Il sifuncolo cirtocoanitico è espanso all’interno delle camere e a
contatto del lato ventrale della conchiglia. I collaretti settali
sono corti e subortocoanitici. Gli anelli di connessione sono cirtocoanitici e
molto espansi nelle camere; il loro diametro massimo è di
circa
Distribuzione - Siluriano superiore della Boemia (Barrande, 1866),
Sardegna sud-orientale (presente tesi).
Collocazione -
collezione paleontologica del Dipartimento di Scienze della Terra, cat. n. 24161 e 24172
Materiale studiato – 2 esemplari.
CENNI
sull’ ECOLOGIA DEI CEFALOPODI
Per
tracciare un quadro chiaro dell’ecologia dei Cefalopodi, occorre anzitutto
definire cosa si intende per cefalopodi. Nella classe Cephalopoda includiamo calamari, polpi e
seppie. Il Nautilus attuale è
inserito tassonomicamente nei cefalopodi nautiloidi, ma è necessario tener
presente che questo fossile vivente è distinto dal punto di vista biologico
dagli altri taxa appartenenti a questa sottoclasse (Nautiloidea).
La
malacologia non ha ancora risolto il problema della relazione fra i cefalopodi
e l’ipotetico mollusco ancestrale provvisto di una
conchiglia conica, una massa viscerale ciliata e di un piede muscolare. Si
suppone che questo mollusco erbivoro pascolasse su
fondali marini di limitata profondità. I più importanti caratteri evolutivi che
hanno differenziato i cefalopodi dagli altri molluschi
sono lo sviluppo di un sistema nervoso gangliare molto raffinato e cefalizzato,
la circolazione chiusa, il moto “a getto”, le ghiandole dell’inchiostro, la
regolazione del galleggiamento e le grandi dimensioni. La
conchiglia è diventata interna ed è in seguito stata persa da molti gruppi; le
pinne si sono evolute per aiutare la locomozione nel mezzo acquatico; i
tentacoli (otto o dieci) si sono specializzati nella cattura delle prede.
L’ecologia del Nautilus attuale è ancora poco conosciuta, soprattutto a causa
delle grandi profondità che predilige. Un singolo esemplare copre infatti grandi spostamenti in senso verticale e spostamenti
molto più brevi in senso orizzontale. Fino al termine del Miocene i nautiloidi
erano distribuiti praticamente in tutti gli oceani,
mentre oggi occupano una zona molto ristretta dell’oceano indo-pacifico,
compresa fra le isole Fiji e l’Australia. Solitamente vive nei pressi di
barriere coralline, ad una profondità minima di
Le massime
profondità di implosione ottenute sperimentalmente
sono dell’ordine di
Movimento verticale
Il Nautilus è un eccellente nuotatore
capace di coprire relativamente grandi distanze. Siccome durante la notte si
osservano esemplari di Nautilus in
acque superficiali dove non se ne osservano durante il
giorno, si pensa che questo animale sia capace di compiere significativi
spostamenti giornalieri in senso verticale. Sperimentalmente si è potuto
riscontrare che la massima velocità di ascesa nella
colonna d’acqua è di
È quindi
chiaro che il nuoto gioca un ruolo importante nella vita del Nautilus. Come gran parte dei
cefalopodi, il Nautilus nuota usando
la testa per direzionare il moto. L’involuzione della
conchiglia conferisce a quest’ultima uno dei disegni più efficaci in termini di efficienza natatoria. Poiché la
conchiglia del Nautilus è involuta e la camera d’abitazione è breve, il centro
di gravità della conchiglia è posto nella camera d’abitazione, mentre il centro
di galleggiamento è esattamente sulla verticale del baricentro.
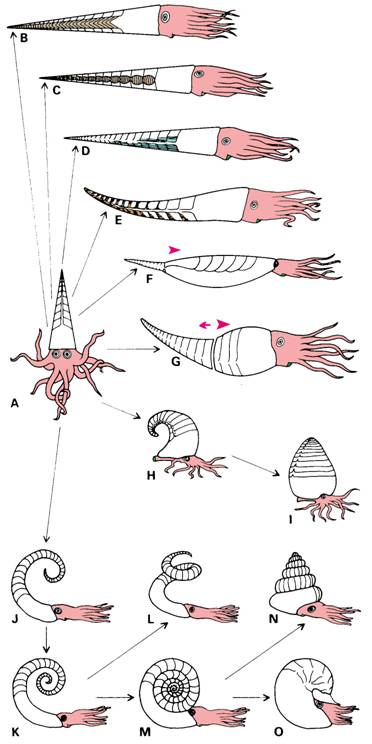
Per quanto riguarda l’assetto delle conchiglie fossili, è stata proposta recentemente una teoria da Cheetham & Powell (1987), che riportiamo di seguito.
Assumendo
come ipotesi un assetto di vita orizzontale per i cefalopodi appartenenti al
necton, quindi liberi natanti, e partendo da una forma diritta (ortocona)
semplice (A) possiamo rilevare dai fossili diverse strategie per ottenere un
aumento di peso nella parte apicale del guscio camerata che altrimenti
tenderebbe ad un galleggiamento positivo variandone
l’assetto, tendendo a portare l’animale con l’apice verso l’alto. Nei
Nautiloidi provvisti di guscio esterno, un aumento di peso apicale può essere
ottenuto con 1) endoconi (Endoceratoidea, B), con complessi depositi annulari
(Actinoceratoidea C) o depositi camerali (vari
Nautiloidea, E). 2) La parte camerata del fragmocono può essere ridotta o
giacere sopra le parti molli con la camera d’abitazione la cui apertura si
adatta per mantenere l’equilibrio (vari Nautiloidea, H-I). 3) Il liquido può
essere trat-tenuto nelle camere per impedirne l’effetto di galleggiamento, D).
4) Le camere possono estendersi oltre le parti molli, così da ridurne l’ effetto di galleggiamento, F). 5) Una parte di fragmocono
può essere abbandonata per impedirne l’effetto galleggiante negli esemplari
adulti (troncatura del guscio, G). 6) La parte camerata può es-sere avvolta in
modo da trovarsi sopra le parti molli (vari Nautiloidea e la maggior parte
degli Ammonoidea J-O).
Non
tutti gli autori concordano col fenomeno della troncatura del fragmocono. Dzik,
1984, discute questo ipotetico fenomeno e lo nega
adducendo che mai nessuna parte “lasciata” da cefalopodi è parte del record
fossile e che le parti abbandonate dei Discosorida (G) possono benissimo essere
spiegate da un fattore meccanico di rottura degli esemplari, durante la loro
raccolta.
Fig. - 1 rappresentazione schematica del
mantenimento dell’equilibrio (da Cheetham & Rowell)
 CENNI DI
PALEOECOLOGIA
CENNI DI
PALEOECOLOGIA
Ci
occuperemo, in questa breve trattazione, degli aspetti paleoecologici relativi
soltanto agli ordini descritti nella parte sistematica.
Ordine Orthocerida
Quasi tutti i rappresentanti di questo gruppo hanno conchiglie ortocona
longicone con un sifuncolo molto piccolo. Sono sempre presenti
voluminosi depositi camerali e sifuncolari nella parte apicale, mentre nella parte ventrale si trovano strutture laminate. Le colorazioni
del guscio sono generalmente limitate alla parte dorsale. La quantità di
depositi progressivamente inferiori verso l’apertura, permette ai rappresentanti
di questo gruppo di mantenere un equilibrio idrostatico orizzontale. La
maggiore concentrazione di depositi camerali nella parte ventrale permette di
abbassare il centro di gravità migliorando la stabilità. In alcuni casi
l’animale provvedeva alla troncatura della conchiglia (non tutti gli autori , ad es. Dzik, 1984, sono concordi ad accettare questa
ipotesi) per ottenere un migliore equilibrio. Si può concludere
che gli Orthocerida fossero nuotatori attivi.
Ordine Oncocerida
In questo ordine sono raggruppate principalmente forme
cirtocone brevicone, ma anche forme ortocona longicone, girocone, torticene,
serpenticone e nautilicone. Negli esemplari adulti si osservano grandi
modificazioni nella forma dell’apertura. Sia il sifuncolo che le camere sono privi di depositi mineralizzati di significativa
entità. Le dimensioni del fragmoconi e della camera d’abitazione è variabile,
ma risultano essere piccole nella maggioranza delle forme. La conchiglia è
spessa e pesante, caratteristiche che non facilitano
certo il nuoto attivo. Per questa ragione gli Oncocerida sono stati
classificati come organismi bentonici che vivevano sui fondali spostandosi
anche con l’aiuto dei tentacoli. È tuttavia presente il seno iponomico, che
indica la presenza dell’imbuto come organo di propulsione, tanto che potrebbe
essere possibile supporre un tipo di vita necto-bentonico. Le ornamentazioni
colorate delle forme brevicone sono distribuite su tutta la superficie della
conchiglia, suggerendo che l’asse fosse inclinato; lo
sviluppo di una curvatura moderata della conchiglia avrebbe permesso una
disposizione orizzontale dell’imbuto. Quindi, le forme
cirtocone con un fragmoconi piuttosto largo potrebbero essere considerate come
buoni nuotatori, nonostante il tipo di conchiglia scarsamente idrodinamico. Si
pongono particolari problemi nel momento in cui si considerano le forme con
aperture fortemente “contratte”, che hanno generalmente un fragmoconi piccolo.
In questo caso potrebbe trattarsi di forme bentoniche, anche se la presenza del
seno iponomico indica la capacità di propulsione.
IMPLICAZIONI
PALEONTOLOGICHE
La maggiore
“invenzione” evolutiva compiuta dai nautiloidi che li ha
affrancati dai fondali marini è stata il fragmocono. Sicuramente sono esistiti
predatori marini antecedenti ai nautiloidi, come prova la fauna del Burgess-Shales
risalente al Cambiano medio. I nautiloidi, tuttavia, essendo comparsi circa 450
milioni di anni fa (nel Cambriano), sono stati i primi
grandi predatori capaci di attaccare anche prede provviste di scheletro.
Probabilmente, il passo più difficile nell’evoluzione di un fragmocono
funzionale, è rappresentato dal sifuncolo. Yochelson et al. (1973) hanno
proposto un cammino evolutivo portante ad un fragmocono funzionale pensando che
il cefalopode ancestrale avesse una serie di setti
chiusi nella parte apicale della conchiglia. Il tessuto dietro alla parte
viscerale rimaneva all’interno di queste parti, attraverso un’apertura
centrale, formando così il primo elemento del sifuncolo. Con la chiusura del sifuncolo
e l’evoluzione di un epitelio pompante si sarebbe originato il fragmocono. A
questo processo sarebbe seguita la formazione dei tentacoli e della regione
cefalica del cefalopode attraverso la fusione della regione cefalica e del
piede del mollusco ancestrale. Tuttavia, non esiste
una prova fossile della validità o della falsità di questa teoria. Dzik (1981),
studiando i cefalopodi del Paleozoico inferiore, ha ipotizzato che l’antenato
dei cefalopodi fosse da ricercare tra i monoplacofori cambriani.
Diversificazione ed
evoluzione dei nautiloidi
Donovan
(1964), Teichert (1967) e Dzik (1984) si sono occupati dell’evoluzione e della
diversificazione dei cefalopodi. Fino a poco tempo fa, la teoria più seguita
era quella secondo cui i cefalopodi si fossero differenziati
nell’Ordoviciano inferiore, ma recenti scoperte (Chen & Teichert, 1983)
hanno dimostrato che la radiazione di questo gruppo è iniziata già nel Cambiano
superiore. Si pensa che gruppo ancestrale da cui si
sarebbero evoluti gli altri nautiloidi sia quello degli Ellesmoceratina, un
piccolo gruppo costituito da specie con conchiglie da leggermente curve a
diritte. Gli ellesmoceratidi non avevano depositi camerali a fare da
contrappeso e quindi la loro posizione di vita sarebbe stata prevalentemente
verticale fino a che non furono in grado di posizionare
del liquido nelle parti più vecchie del fragmocono. Dagli ellesmoceratidi si
sarebbero poi radiati una gran quantità di gruppi di nautiloidi paleozoici. Tra
questi si possono distinguere due linee evolutive per quanto riguarda la forma
della conchiglia, la prima delle quali porta a conchiglie ortocone e diritte.
Questa particolare forma della conchiglia, tuttavia, avrebbe
richiesto la formazione di massicci depositi camerali per far mantenere un assetto
di vita orizzontale agli organismi. L’altro trend
morfologico ha portato a conchiglie planispirali. Le forme con depositi
camerali, collocati sia nelle camere sia nel sifuncolo, avrebbero dovuto pagare
un prezzo significativo in termini di mobilità e di
spesa energetica per mantenere l’assetto orizzontale. Infatti, i depositi
camerali avrebbero occupato significative parti del
fragmocono ed avrebbero quindi richiesto lo sviluppo di fragmoconi più
voluminosi, a parità di volume della camera d’abitazione, per mantenere un
galleggiamento neutro. Tuttavia, un nautiloidi provvisto di questi depositi
camerali avrebbe subito un aumento di massa, che, dal punto di vista
energetico, avrebbe reso più dispendiosi tutti i
movimenti rispetto ad un esemplare di volume equivalente e massa minore. Alcuni
degli ortoconi paleozoici raggiungevano dimensioni ragguardevoli e non sarebbero stati molto efficienti nell’inseguire le prede.
Il modo in
cui vengono deposti i depositi camerali è stato
oggetto di molte dispute. Alcuni autori hanno ipotizzato l’esistenza di un
tessuto vivente all’interno delle camere in grado di produrre i depositi
camerali. In molti casi non si riscontrano tracce della vascolarizzazione
necessaria per un tale tessuto. Altri autori hanno ipotizzato che i depositi
camerali non siano prodotti durante la vita del
nautiloidi. Alcuni ritrovamenti di Crick (1982) hanno dimostrato l’infondatezza
di questa idea. Fischer (1969) ha pensato che i
depositi camerali precipitassero direttamente dal liquido camerale,
processo di cui è stata dimostrata la possibilità.
La forma
delle conchiglie dei generi del Paleozoico mostra un
graduale cambiamento da faune dominate da forme diritte o curve a faune
dominate da forme planispirali. Al termine del Paleozoico i nautiloidi andarono
incontro ad una drastica riduzione.
Lo studio
dei setti, della curvatura, degli anelli di connessione e dello spessore delle
forme fossili può fornire informazioni sulla profondità a cui queste ultime
vivevano. Queste strutture, infatti, rappresentano fattori limitanti rispetto
alla profondità in quanto possono implodere se sottoposte a pressioni troppo
elevate. Westermann concluse che forme con setti ravvicinati, pareti della
conchiglia, pareti settali e anelli di connessione
spessi dovevano essere più resistenti ad alte pressioni. Di questi caratteri,
quelli più utili sono lo spessore e le dimensioni degli anelli di connessione,
perché questi sono gli elementi a più alto rischio di implosione,
in quanto separano due zone sottoposte a pressioni molto diverse (le camere
hanno una pressione sempre inferiore a 1 atmosfere, mentre la parte interna
dell’anello può essere sottoposta a pressioni anche di 60 atmosfere).
Westermann propose (1971) l’”indice relativo di resistenza del sifuncolo”,
ottenuto dalla formula 100 (h/r), in cui h è lo
spessore del sifuncolo e r è il suo raggio interno. Siccome
il Nautilus attuale implode ad una profondità di circa
CONCLUSIONI
La fauna
studiata è costituita da 14 esemplari appartenenti a sei taxa che non erano mai stati rinvenuti
nella regione presa in esame. Fra questi, il più frequente è risultato essere Michelinoceras cf. michelini (5 records), seguito da Michelinoceras cf. currens, Kopaninoceras
cf. thyrsus,
Columenoceras cf. grande, Temperoceras cf. temperans e Oonoceras
aff. potens con 2 records ciascuno.
BIBLIOGRAFIA
Barrande J.,
1865-74. Sistéme Silurien du centre de
Boardman R. S., Cheetham A.
H., Rowell A. J., 1987. Fossil
Invertebrates, Blackwell Scientific Publications.
Carmignani
L., Gattiglio M., Maxia M., Oggiano G., Pertusati P. C., 1986.
The geology of
Gerrei. Paleozoic
basement of
Corradini
C., Ferretti A., Serpagli E., 1998. the
Silurian and Devonian sequence in SE Sardinia, Giorn. Geol, ser. 3a ser.,
vol. 60, spec. Issue, ECOS VII-Sardinia Guide-Book, pp. 71-74.
Corradini
C., Leone F., Loi A., Serpagli E., 2001. Conodont stratigraphy of
a highly tectonised Silurian-Devonian section in the San Basilio area (SE
Sardini,
Dzik J., 1984. Phylogeny of the
Nautiloidea. Palaeontologia Polonica, n. 45, pp. 3-203, 47
pls., Warszawa – Krakow.
Gnoli M., 1987. Revision and autoecological
remarks of the species Columenoceras grande (Meneghini, 1857) (Nautiloidea,
Orthocerida). Boll. Soc. Paleont. It., 26 (3),
245-250.
Gnoli M., 1990. New evidence for faunal
links between Sardinia and
Lenoe F., 1998. Post
Sarrabese Ordivician sequence in SE Sardinia, Giorn. Geol, ser. 3a ser., vol. 60, spec. Issue, ECOS VII-Sardinia Guide-Book,
pp. 63-69.
Meneghini G., 1857.
Paléontologie de l’Ile de Sardaigne (in
Naud G.,
1982. Schema stratigrafico strutturale del Paleozoico della
zona di S. Basilio (Sardegna sud-orientale) – Guida alla Geologia del
Paleozoico Sardo. Guide
Geologiche Regionali. Soc. Geol. It., 1982: 109-115.
Saunders W. B. & Landman
N. H., 1987. NAUTILUS
the biology and Paleobiology of a living fossil. Plenum Press.
Serpagli E. & Gnoli M., 1977. Upper Silurian Cephalopods from
Summers W. C., 1983. Phyisiological and Trophic Ecology
of Cephalopods, in Russell-Hunter
W. D. (Eds.), 1983. The Mollusca volume
6 Ecology. Academic Press Inc.
Sweet W. C., 1964. Nautiloidea-Oncocerida in
R.C. Moore (ed.), Treatise on invertebrate Paleontology. – Soc. Geol.
Am. And Univ.
Ward P. D., 1987. The natural history of
Nautilus. Allen & Unwin.
TAVOLA 1
Figura 1: Michelinoceras cf. currens x 1
Figura 2: Michelinoceras cf. michelini x 1,2
Figura 3: Kopaninoceras cf. Thyrsus x 1

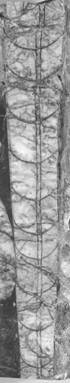

1 2 3
TAVOLA 2
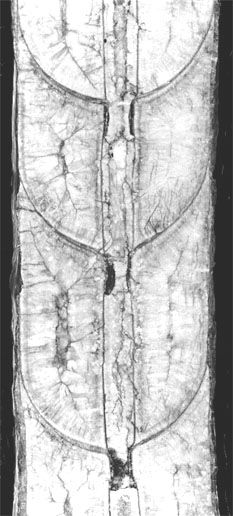
Figura 1: Columenoceras cf. grande x 2
Figura 2: Temperoceras cf. temperans x 1

1 2
TAVOLA 3
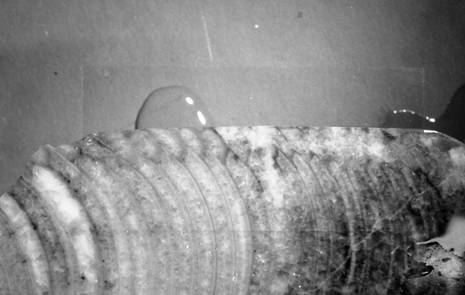
Figura 1: Oonoceras aff. potens x 2
Figura 2: Oonoceras aff. Potens, particolare del sifuncolo x 4

1