DINOSAURI, I
DOMINATORI DEL MESOZOICO
SISTEMATICA,
EVOLUZIONE E
SCOMPARSA
DEI PIÚ GRANDI RETTILI
Ad Achille, compianto
maestro
di Paleontologia, e a
Maurizio,
mio mentore.
I DINOSAURI
I Dinosauri rappresentano non solo le forme fossili più
popolarmente conosciute e più comuni nell’immaginario collettivo, ma anche
quelle che hanno dominato l’ambiente terrestre dal Triassico superiore alla
fine del Cretaceo, cioè da
Popolarmente sono riuniti spesso in un singolo
gruppo, ma occorre distinguerli in due ordini, Saurischia ed Ornitischia,
i cui rappresentanti non sono apparentati tra loro più di quanto lo siano
Pterosauri e Coccodrilli. Questa separazione, un tempo basata prevalentemente
sulla forma del cinto pelvico, triradiato nei Saurischi e tetraradiato negli
Ornitischi, trova oggi le sue ragioni d’essere in un’origine separata dei due
gruppi.
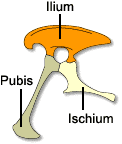
Cinto pelvico di Ceratosaurus, tipico Cinto pelvico di Thescelosaurus,
un
rappresentante dei Saurischi tipico Ornitischio.
Il motivo iniziale del successo: la camminata
Tutti gli Arcosauri, gruppo a cui appartiene gran parte dei Dinosauri e i cui unici rappresentanti attuali sono i Loricati (coccodrilli), tendono a portare gli arti in posizione più sottostante al corpo rispetto a tutti gli altri Rettili e a camminare con un movimento dell’arto “avanti-indietro” e con l’arto tendente al verticale. Nei Tecodonti questa camminata, definita «parasagittale», è raggiunta anche grazie ad un leggero disassamento della testa dell’omero/femore, ma per lo più grazie ad una curvatura del femore stesso, che è concavo verso l’interno. Già nei primi rappresentanti dei Dinosauri, l’articolazione posteriore tra femore e cinto pelvico, è molto specializzata e la camminata è effettuata con gli arti molto più verticali che nei precedenti Rettili. La testa del femore è molto più disassata e vergente verso l’interno del corpo, e la cavità acetabolare è molto profonda, in modo da sopportare meglio il peso del corpo. Contemporaneamente, nella parte superiore del femore, si sviluppano delle tuberosità, dette piccolo e grande trocantere, che fungono da inserzione per i due muscoli ileofemorali. Nella cavità acetabolare, inoltre, si apre una fenestrazione che permette il passaggio ai vasi sanguigni, al tessuto nervoso e ai fasci di fibre che legano il femore al cinto e, indirettamente, alla colonna vertebrale. Nelle forme bipedi la coda è sempre utilizzata come bilanciere.
Ordine Saurischia
(Triassico
superiore-Cretaceo superiore)
Sono caratterizzati da un bacino di tipo triradiato, più primitivo, e ciò li rende simili ai Tecodonti (arcosauri ancestrali). In questo gruppo le forme bipedi carnivore sono prevalenti su quelle quadrupedi, che raggiungono però le massime dimensioni.
Da un punto di vista
tassonomico, vengono distinti in due Sottordini:
Theropoda forme primitive, bipedi, carnivore.
A loro volta si dividono in Coelurosauria e Carnosauria. Questa divisione non ha base
scientifica, ma serve a distinguere le forme di dimensioni relativamente
piccole
e corritrici (Coelurosauri) dalle forme di grandi dimensioni (Carnosauri). Per questioni pratiche conserviamo questa divisione, anche se spesso si trovano esemplari presentanti caratteri intermedi tra i due gruppi.
Sauropoda forme
specializzate, semibipedi o quadrupedi, erbivore. Anche in questo caso
si ricorre ad una classificazione artificiale di comodo, suddividendo il
Sottordine in Prosauropoda, i cui membri rappresentano
forme di passaggio fra Theropoda e Sauropoda, e Sauropoda strictu sensu.
I Theropodi primitivi
La prima forma ritenuta appartenente ai Dinosauri Theropodi per i caratteri del cinto pelvico, dell’arto posteriore e dei denti, è Staurikosaurus, proveniente dal Trias medio del Sud del Brasile. Di questa forma sono conosciuti soltanto la colonna vertebrale, il cinto pelvico, la mandibola, il femore, il radio e l’ulna. Herrerasaurus, del Trias superiore del Sud America, è conosciuto leggermente meglio. Queste due forme presentano caratteri che non si discostano da quelle successive; occorre mettere in particolare evidenza la tendenza alla tridattilia nell’arto superiore, per ablazione del quinto dito e riduzione del primo, e all’allungamento dei metatarsali, caratteri che si ritroveranno sviluppati successivamente.
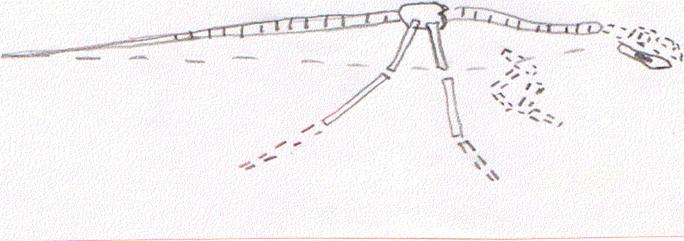
Ricostruzione di Starikosaurus, il più antico
dinosauro conosciuto, proveniente dal Triassico medio del Sud America.
Lunghezza totale
Theropodi Coelurosauri
Sono caratterizzati dagli arti posteriori allungati,
con grande sviluppo dei metatarsali, fatto che denota la loro attitudine alla
corsa. Il cranio è in genere molto piccolo, con grandi orbite e fenestrazioni
molto arretrate. Sono forme comuni dal Trias superiore al Cretaceo terminale. Coelophisis
è una forma tipica del Trias superiore del Nord America, ed era lungo circa

Coelophysis, piccolo
teropode preveniente dal Triassico superiore degli Stati Uniti occidentali.
Durante il Giurassico superiore è presente in Europa
Compsognathus, il più piccolo dinosauro conosciuto, che probabilmente
non doveva pesare più di 3 chili e che presenta l’arto anteriore provvisto di
due sole dita. Un gruppo molto interessante, frequente nel Cretaceo del Nord
America e dell’Asia, è quello degli Ornitomimidi (Ornithomimus, Struthiomimus
ed Oviraptor ne sono gli esponenti principali), che avevano le
dimensioni e la struttura corporea degli attuali struzzi e che si distinguono
dagli altri Dinosauri per l’assenza totale di dentatura. Tutte queste forme rientrano
chiaramente nel gruppo formale dei Coelurosauridi, ma alcune forme, tra cui
ricordiamo in particolare Deinonychus del Cretaceo inferiore del
Montana, presentano caratteri quali tronco e collo accorciati, cranio molto
sviluppato e metatarsali non molto allungati, che li collocano a metà strada
tra Coelurosauri e Carnosauri.

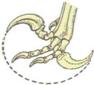
Confronto fra Deinonychus e uomo. Rappresentazione schematica di un artiglio di Deinonychus
Theropodi Carnosauri
Fatta eccezione per le forme precedentemente citate,
tutti gli altri Teropodi appartengono al gruppo dei Dinosauri carnivori, che
hanno espresso i più grandi carnivori mai esistiti. Questi animali potevano
infatti raggiungere i quindici metri di lunghezza e un peso di settanta
quintali. Le forme del Triassico sono ancora oggetto di studio per gli esperti.
Dal Giurassico superiore al Cretaceo superiore (non terminale) Nord America ed
Asia erano popolati dagli Allosauridi. Allosaurus, risalente al
Giurassico superiore del Nord America, è una forma primitiva, con struttura
abbastanza massiccia, con femore e tibia/fibula all’incirca della stessa
lunghezza e metatarsali non molto allungati e parzialmente fusi tra loro.
L’arto anteriore è ridotto e probabilmente non veniva utilizzato nella
camminata. Il cranio è abbastanza allungato e stretto, ed i denti sono
acuminati e rivolti all’indietro. Le fenestrazioni postorbitarie sono grandi,
indice di una potente muscolatura. Un esemplare di Allosaurus poteva
raggiungere dodici metri di lunghezza.
I Tirannosauridi sono i giganti del gruppo, e
si distinguono dai precedenti per un ulteriore accorciamento del torace e del
collo, per gli arti anteriori estremamente ridotti e per un cranio più corto
(anche se superava abbondantemente
Ricostruzione di un esemplare di Tyrannosaurus rex.
Teropodi erbivori: i Sauropoda
Occorre precisare che viene utilizzata, anche in
questo caso, una classificazione di comodo per distinguere le forme primitive
del Trias superiore/Giurassico inferiore (Prosauropoda) da quelle evolute del Giurassico/Cretaceo
(Sauropoda strictu sensu).
Le forme triassiche,
rappresentate da Prosauropoda (o Plateosauria) provengono dai giacimenti
del Triassico superiore tedesco (Plateosaurus), del Nord e Sud America (Mussaurus), Sud Africa (Plateosauravus)
e Cina (Lugengosaurus). Nel Trias superiore, quindi, troviamo forme già
ben differenziate e che occupavano un ampio areale geografico.
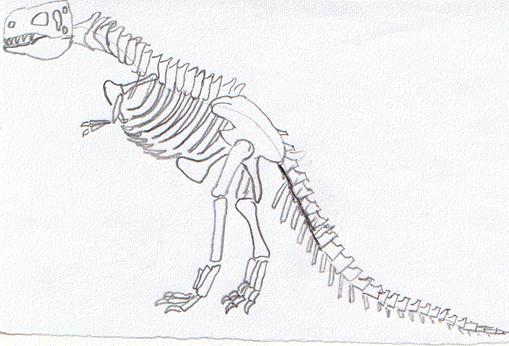
Scheletro di Plateosaurus,
uno dei dinosauri più primitivi, proveniente dal Triassico medio-superiore
della Germania. Misurava fino a
Plateosaurus, lungo fino a sette metri, presentava arti
anteriori ridotti rispetto a quelli posteriori. Il cranio, relativamente
piccolo rispetto al resto del corpo, portava denti guanciali molto numerosi e
differenziati, spesso tricuspidati, mentre i denti centrali erano di forma vagamente
conica. Il collo era moderatamente allungato, a differenza della potente coda.
Il piede mostra quattro dita, per riduzione del quinto, mentre la mano è
completa di cinque dita.
Le origini dei Sauropodi sono estremamente oscure:
per i caratteri del cinto posteriore, li si può avvicinare ai Teropodi più
primitivi del Trias medio (Staurikosaurus ed Herrerasaurus). I
caratteri di primitività si riscontrano anche nella postura: il femore si
discosta di un angolo di circa venticinque gradi dall’asse verticale, mentre
nelle forme più evolute questo angolo è inferiore ai quindici gradi.
I veri Sauropodi, naturale evoluzione dei precedenti
e rappresentati da Diplodocus, Brachiosaurus, Brontosaurus
e Camarasaurus, sono caratterizzati dalle enormi dimensioni: potevano
raggiungere trenta metri di lunghezza e un peso variabile fra le settanta e le
ottanta tonnellate. A causa di queste dimensioni, dovevano essere forzatamente
quadrupedi, con arti solidi portati quasi verticalmente. Questo tipo di arto è
detto arto colonnare a struttura graviportale, con omero e femore più lunghi in
rapporto a tibia e fibula. Questo fenomeno si riscontra anche in alcuni
mammiferi di grandi dimensioni (Proboscidati) e, più in generale, caratterizza
le forme camminatrici.
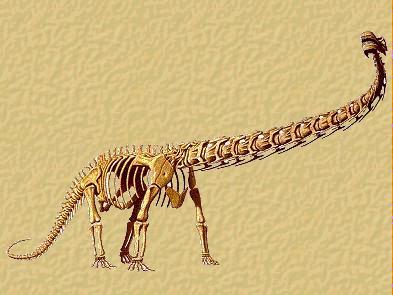
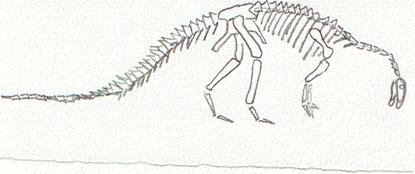
Ricostruzione dello scheletro di un Diplodocus.
Il cranio di queste forme è sempre molto piccolo in
rapporto alle dimensioni corporee: il Diplodocus, che poteva misurare
fino a trenta metri di lunghezza, aveva un cranio lungo “soltanto”
Ordine Ornitischia
(Giurassico inferiore –
Cretaceo superiore)
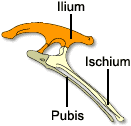
Costituiscono un gruppo molto eterogeneo, e si distinguono dai precedenti soprattutto per il bacino tetraradiato, morfologicamente molto simile a quello degli Uccelli. Il bacino è infatti costituito da un ileo molto espanso, da un ischio rivolto verso la parte posteriore e da un pube espanso sia anteriormente che posteriormente, parallelo all’ischio.
Ricostruzione scheletrica del bacino di un ornitischio.
La grandezza del pube potrebbe essere riconducibile
al bipedismo delle forme primitive, che, essendo erbivore ed avendo quindi una
cavità addominale di grandi dimensioni, necessitavano di una muscolatura assai
efficiente per poter mantenere una camminata di tipo parasagittale. Un altro
carattere comune a tutti gli Ornitischi è la presenza del predentale nella
mandibola, un’ossificazione accessoria priva di dentatura. Per quanto riguarda
le origini, non è mai stata trovata una forma che possa rappresentare il
tramite tra Saurischi ed Ornitischi, in quanto anche le forme primitive
presentano già caratteri distinti.
Gli Ornitischi vengono generalmente divisi in cinque
sottordini:
Ornithopoda forme bipedi primitive del
Triassico superiore-Cretaceo
Pachicephalosauria forme con cranio globoso e
ispessito
(Cretaceo).
Stegosauria forme quadrupedi, con
doppia fila di piastre ossee dorsali (Giurassico
superiore-Cretaceo inferiore).
Ankilosauria forme provviste di corazza
ossea dermica (Giura superiore-Creta superiore).
Ceratopsida forme con cranio terminante
con una specie di becco (Cretaceo superiore).
Sottordine Ornithopoda
La forma più antica
conosciuta proviene dal Triassico superiore argentino: Pisanosaurus,
poco conosciuto a causa della scarsità e incompletezza dei resti rinvenuti, che
mostra già netti caratteri ornitischi, come la presenza del predentale. Molto
meglio conosciuto è Fabrosaurus, del Giurassico basale del Sudafrica.
Questa forma presentava uno scheletro molto leggero e simile a quello dei
primitivi Saurischi quali Coelophysis, con arti anteriori ridotti, che
denotano uno spiccato bipedismo. Il cranio è piccolo ed accorciato, con la
fenestrazione preorbitaria parzialmente occlusa, come in gran parte degli
Ornitischi, dall’espansione dei mascellari. Il premascellare presenta ancora i
denti, che andranno perduti nelle forme successive, mentre nella mandibola è
presente un rudimentale predentale, caratteristico di tutti gli Ornitischi. La
forma dei denti, compressi lateralmente, segnala una dieta di tipo
onnivoro/erbivora. Nel cinto pelvico, l’ischio è molto allungato in senso
antero-posteriore, mentre il sottilissimo pube non presenta alcuna espansione
rivolta anteriormente. La testa del femore non è molto disassata sotto la
colonna vertebrale, fatto che indica che durante la camminata il femore non era
verticale, ma era portato in modo leggermente divaricato.
Altrettanto primitivo è Heterodontosaurus,
contemporaneo di Fabrosaurus, che presenta una dentizione differenziata
e arti anteriori più sviluppati, che gli potevano permettere anche un’andatura
quadrupede. Da queste forme ancestrali si svilupperanno diverse linee
evolutive, che si differenzieranno fino al Campaniano. Per gli Ornitopodi
evoluti esamineremo in questa sede soltanto le tre linee filetiche principali:
Ipsilofodonti, Iguanodonti e Adrosauri.
Gli Ipsilofodonti del Giurassico
superiore-Cretaceo rappresentano il gruppo più conservativo all’interno degli
Ornitopodi, ripetendo, in modo leggermente più evoluto, i caratteri visti in Fabrosaurus,
da cui si differenziano per il pube espanso anche anteriormente e per l’assenza
di dentatura sul premascellare. Raggiungono inoltre maggiori dimensioni, fino a
cinque metri di lunghezza. Sono presenti dal Giurassico superiore in Europa,
Nord America ed Australia. Il genere più caratteristico e rappresentativo di
questo gruppo è Hypsilofodon.
Gli Iguanodonti sono caratterizzati da
grandi dimensioni e dalla struttura graviportale degli arti posteriori;
probabilmente si sono evoluti da antenati Ipsilofodonti. Potevano avere
stazione tanto bipede quanto quadrupede. Camptosaurus è una forma
abbastanza primitiva proveniente dal Giurassico superiore del Nord America. Ha
un cranio allungato, con predentale e premascellari privi di denti. I denti
guanciali, compressi lateralmente e taglienti, dovevano essere adatti a
strappare i vegetali. I robusti arti presentano una struttura quasi
graviportale, con quelli posteriori più sviluppati di quelli anteriori. Poteva
raggiungere una lunghezza di sei metri. Tipico rappresentante di questo gruppo
è Iguanodon, del Cretaceo inferiore inglese, che è stato in
assoluto il primo dinosauro ad essere descritto.
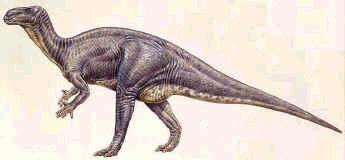
Ricostruzione di Iguanodon.
Gli Adrosauri,
o “dinosauri dal becco d’anatra”, sono forme comuni nel Cretaceo superiore del
Nord America, Asia, Europa e Sud America. Strettamente collegati al mezzo
acqueo, presentano arti posteriori molto più sviluppati di quelli anteriori, e
un piede allargato, quasi palmato. Caratteristica del cranio di queste forme è
la grande espansione delle narici interne. I denti erano costituiti da piastre
dentarie. Gli Adrosauri si estinsero al limite fra Cretaceo e Terziario. Le
forme più rappresentative di questo gruppo sono Corythosaurus, Anatosaurus
e Limnosaurus.
Sottordine Pachicephalosauria
(Cretaceo)
Si tratta di un piccolo gruppo
di antiche origini, che probabilmente rappresenta una linea filetica separatasi
dagli Ornitopodi. Sono forme relativamente piccole, bipedi, la cui
caratteristica principale è quella di avere un cranio globulare, dovuto ad un
ispessimento dei parietali e dei frontali. Yaverlandia, la forma più
antica conosciuta, viene dal Cretaceo inferiore inglese, ma esemplari
appartenenti a questo gruppo sono stati ritrovati anche in Nord America ed in
Asia.
Sottordine Stegosauria
(Giurassico medio-Cretaceo
superiore)
Sono forme che hanno
conservato la protezione ossea dermica tipica di molti Tecodonti. La forma più
conosciuta e rappresentativa del gruppo, del Giurassico superiore del Nord
America, è Stegosaurus, che presenta una doppia fila di piastre osse
mobili assai vascolarizzate, situate sopra la colonna vertebrale ed orientate
perpendicolarmente ad essa, che probabilmente fungevano da “pannelli solari”.
Forme analoghe sono Omosaurus (Europa) e Kentrosaurus
(Africa).

Ricostruzione di un esemplare di Stegosaurus.
La forma ancestrale di questo gruppo può essere
individuata in Scutellosaurus, che era provvisto di piastre dermali in
corrispondenza delle vertebre.
Sottordine Ankylosauria
(Giurassico
superiore-Cretaceo superiore)
Possono essere considerate forme corazzate, in
quanto l’intero tronco era rivestito da piastre ossee dermiche. Anche il cranio
è ricoperto da piastre ossee saldate alle ossa craniche. Queste piastre erano
disposte in “strisce” articolate fra loro. La coda in alcune forme era munita
di espansioni ossee terminali, analoghe alle spine che si trovano sulla coda di
alcuni Stegosauri. Si tratta sempre di forme quadrupedi, con arti tozzi e
corti. La testa del femore non è molto disassata, quindi la loro camminata è
intermedia tra la reptazione e la camminata parasagittale. La dentatura è
sempre poco sviluppata e, in qualche caso, assente. Le forme più comuni sono Ankylosaurus
(Cretaceo superiore dell’Alberta), lungo più di quattro metri; Scolosaurus,
provvisto di spine su tutto il corpo, Acanthopholis (Cretaceo superiore
inglese) e Struthiosaurus (Cretaceo superiore francese e della
Transilvania).
Sottordine Ceratopsia
(Cretaceo)
Sono un gruppo che offre un esempio di sempre
maggiore specializzazione. Gli Psittacosauri, le forme più primitive, compaiono
nel Cretaceo inferiore cinese, e presentano predentali, premascellari e rostro
conformati a simulare un becco di pappagallo. Psittacosaurus presenta
un’andatura tipicamente bipede, con arto anteriore moderatamente ridotto. Il
fatto che la parte distale dell’arto posteriore sia più lunga di quella
prossimale, denota l’attitudine alla corsa. Probabilmente i Ceratopsidi
derivano dai Protoceratopsidi del Cretaceo superiore, come Protoceratops,
forma cinese di non grandi dimensioni, nel cui cranio si assiste ad uno
spostamento all’indietro degli squamosi e dei parietali, che vanno a costituire
un collaretto che ricopre le vertebre cervicali e che funge da ampia base di
ancoraggio per i muscoli che chiudono le mandibole. Il grande successo dei
Ceratopsidi all’inizio del Cretaceo può essere dovuto all’estrema
specializzazione ed efficienza dell’apparato di assunzione del cibo. Nelle
forme più evolute compaiono anche corna ossee: dapprima un solo corno formato
da un’espansione dei nasali (Monoclonius), e nelle forme finali tre
corna formate da postorbitali e parietali (Triceratops). Le forme evolute di Ceratopsidi sono
esclusive del Nord America, le forme primitive di Psittacosauri si ritrovano
soltanto in Asia, mentre i Protoceratopsidi si ritrovano in entrambi i continenti.
Il mistero dell’estinzione
La fine del Cretaceo
(66.5 o 65.6 milioni di anni fa secondo la cronologia assoluta) si caratterizza
per la scomparsa di molte forme terrestri, fra cui Dinosauri (Saurischi ed
Ornitischi) e Pterosauri, mentre in ambiente marino si estinsero Plesiosauri e
Mosasauri. Pur tuttavia, i dinosauri non furono gli unici gruppi a subire
questo passaggio (anche Marsupiali e Multitubercolati andarono incontro a una
forte riduzione numerica). L’interpretazione di questa crisi ha da sempre
stimolato i geologi e anche scienziati che hanno considerato il fenomeno
soltanto dal punto di vista fisico, trascurando completamente le componenti
biologiche. Per questo è ancora accesissima una sorta di lotta tra i “gradualisti”
e i “catastrofisti”.
LA GRANDE
CATASTROFE
L’ipotesi catastrofica ha
preso piede dopo i lavori di Alvarez et alii
durante gli anni ’80. base di questa ipotesi è la constatazione che nelle rocce
sedimentarie la concentrazione di Iridio è estremamente bassa (0.3 parti
per blione), mentre è leggermente più alta nelle rocce ignee ricche in ferro,
in cui l’Iridio è solubile. Assai più ricchi di Iridio sono i meteoriti (0.5
parti per milione). In molte successioni stratigrafiche, il passaggio tra
Cretaceo e Terziario è marcato da uno strato, generalmente molto sottile, in
cui si ha un notevole arricchimento in Iridio, fino a 160 volte la norma.
Questo strato, segnalato per la prima volta in località Bottaccione, nei pressi
di Gubbio, venne ritrovato in molte altre località, dal Portogallo ad Israele,
dal Sud America alla Nuova Zelanda. Su questa base, Alvarez (che è un fisico)
ipotizzò che un meteorite di grandi dimensioni (almeno dieci chilometri di
diametro) fosse stato catturato dalla gravità terrestre e avesse orbitato per un
certo tempo attorno alla Terra. Il contatto con l’atmosfera l’attrito avrebbe
provocato una disgregazione almeno parziale del corpo celeste. L’impatto,
avvenuto presumibilmente nella zona dell’attuale Yucatan, avrebbe
sollevato una nuvola di polvere densa che avrebbe oscurato
L’IPOTESI GRADUALISTA
A – Gli avvenimenti che precedono la crisi
Durante l’ultima parte
del Cretaceo, l’estensione dei continenti era andata riducendosi a causa di un
innalzamento del livello del mare. In queste condizioni di circolazione delle
correnti marine e con la frammentazione dei continenti, il clima era
probabilmente temperato caldo ed abbastanza uniforme, con bassa
stagionalità. Verso la fine del Cretaceo, queste condizioni iniziarono a mutare
a causa dell’attività tettonica: le Americhe si stavano allontanando da Europa
e Africa, mentre l’allontanamento della Groenlandia da America ed Europa,
provocò il riversarsi di acque fredde verso basse e medie latitudini. La circolazione
delle acque marine iniziò ad organizzarsi in modo più simile a quello attuale,
con un conseguente raffreddamento generale, indicato dai cambiamenti delle
flore. In Wyoming si passò da temperature annuali medie superiori ai venti
gradi centigradi nel Maastrichtiano, a temperature di poco superiori ai dieci
gradi nella parte iniziale del Terziario. Questo raffreddamento portò
all’abbassamento del livello marino e alla scomparsa del mare interno nord
americano. Esiste poi il fatto innegabile delle polveri vulcaniche
originate dallo scontro far Asia e India, come anche esistono i segni
dell’impatto con un corpo celeste, ma risulta assai difficile valutare gli
effetti di questi eventi sulla vita.
B – I VERTEBRATI TERRESTRI COINVOLTI
Approssimativamente, i Vertebrati terrestri viventi
al passaggio fra Cretaceo e Terziario erano i Dinosauri (Saurischi ed
Ornitischi), i Crocodylia, i pochi rettili volanti, un gran numero di
Lepidosauri, generalmente di piccole dimensioni, e due Sottoclassi dei
Mammiferi: gli Alloteri, rappresentati dai Multitubercolati (oggi completamente
estinti), e i Therii, rappresentati da Marsupiali e Placentati, tutti di
piccole dimensioni. Per quanto concerne i “Dinosauri”, la loro massima
differenziazione si era verificata nel Campaniano, cioè circa dieci milioni di
anni prima della grande crisi. Delle 24 famiglie di Dinosauri esistenti nel
Campaniano, già nel Maastrichtiano superiore, ne sopravvivono soltanto otto.
Una considerazione analoga deve essere fatta per i Rettili volanti, di cui nel
Maastrichtiano ne sopravvive una sola famiglia, composta da tre soli generi di
grandi dimensioni, che si estingueranno alla fine del Cretaceo. Discorso a
parte meritano invece i Lepidosauri, che si erano differenziati nel Giurassico
ed erano rappresentati da un gran numero di Famiglie, con esemplari
generalmente di dimensioni medio/piccole. Passarono il famigerato limite K/T
apparentemente senza problemi, ad eccezione dei Mosasauri, Rettili marini.
Dalla crisi di fine Cretaceo non furono interessati nemmeno i Testudinati.
È doveroso aprire una parentesi sul caso dei
Mammiferi, che durante il Maastrichtiano erano in grande espansione. I
Multitubercolati, rappresentati nel Cretaceo da undici generi, nel Paleogene
iniziale consteranno di soli sette generi. Anche i Marsupiali subiscono una
profonda crisi: alla fine del Cretaceo ne esistevano tre o quattro famiglie,
mentre nel Paleocene è presente soltanto
C
– CONTEMPORANEITÀ DELLE ESTINZIONI
Per determinare la natura dell’evento che
caratterizza la fine del Mesozoico (K/T), è necessario sapere per quanto tempo
questo evento è durato e conoscere anche la sincroneità nei vari ambienti. Il
criterio paleobiologico, che stabilisce l’inizio dell’era Terziaria sulla base
della comparsa di una o più specie tipicamente terziarie, non è sempre
applicabile con certezza. Una correlazione abbastanza esatta è possibile quando
si opera in ambiente marino, ma è difficile correlare tra loro sedimenti
marini, d’acqua dolce e continentali, specialmente se sono separati da grandi
distanze. Anche le datazioni radiometriche sono, per un evento lontano nel
tempo come il K/T, abbastanza imprecise, ammettendo un errore di un milione di
anni in più o in meno. In definitiva, il criterio più sicuro per il
riconoscimento del limite K/T è basarsi sull’anomalo picco di Iridio nei
sedimenti. Per quanto riguarda l’estinzione dei Dinosauri sulla terraferma,
l’unico continente accuratamente studiato è il Nord America. Nel Montana e
nell’Alberta (Canada) la fauna del Campaniano superiore era composta da circa
trenta generi; nel Maastrichtiano superiore (non terminale) ne sopravvivono
soltanto tredici. Anche in altre località si riscontra un impoverimento della
fauna; nell’ultima parte del Maastrichtiano superiore, l’80% dei resti fossili
è rappresentato da Triceratops. Si può ipotizzare che l’ambiente
del Maastrichtiano superiore fosse un ambiente stressato e che Triceratops
potesse essere una forma opportunista. Sarebbe quindi plausibile pensare che
l’estinzione non sia sta improvvisa, ma che vi sia stato piuttosto un lungo
periodo di crisi, culminato con l’estinzione finale. Per quanto riguarda la
contemporaneità di queste estinzioni, occorre tener presente che nel Montana,
in sedimenti fluviali di sicura età paleocenica (in quanto sovrastanti il picco
di Iridio), sono stati ritrovati resti isolati di Dinosauri. Questo fatto non
prova però che i Dinosauri siano vissuti anche in età paleocenica, in quanto i
resti potrebbero essere stati erosi da un deposito del Cretaceo e rideposti in
sedimenti paleocenici. Possiamo soltanto affermare che i Dinosauri hanno avuto
il massimo della loro diversificazione nel Campaniano, che nel Maastrichtiano
presentavano un sempre più basso livello di diversificazione, e che l’ambiente
era “stressato”, con il sopravvento di specie opportuniste.
LA FINE DEI DINOSAURI
I Dinosauri hanno rappresentato, tra i Vertebrati
terrestri, le forme dominanti dal Triassico superiore fino al Cretaceo
superiore, cioè da
Perché i Dinosauri raggiunsero questa posizione di
predominio e per quale ragione si estinsero senza essere entrati in evidente
competizione con altri Vertebrati?
Per quanto riguarda
l’inizi del loro predominio, va detto che ciò avvenne nel Trias superiore, in
corrispondenza con un trend
che portò il clima sulla terraferma ad essere più caldo e secco. Molti dei
cosiddetti “Rettili mammaliani”, più adatti alle zone caldo-umide, erano già
estinti. Per scoprire le ragioni dell’estinzione dei Dinosauri, è necessario
fare alcune considerazioni sulla loro biologia e sugli avvenimenti del Cretaceo
superiore.
Premessa: ECTOTERMIA e ENDOTERMIA
Il metabolismo dei
Mammiferi e degli Uccelli attuali differisce da quello degli altri Vertebrati
per molti caratteri, in quanto permette un’attività fisica più continuata nel
tempo e presenta la fonte primaria del calore corporeo. Il metabolismo
basale si esprime in termini di quantità di ossigeno consumato nell’unità
di tempo per unità di peso quando il corpo è in assoluto riposo. A parità di
peso corporeo, il metabolismo di un mammifero è circa 10 volte superiore
rispetto a quello di un rettile. Sotto sforzo, il metabolismo di un rettile
aumenta da sei a quattordici volte, quello di un mammifero aumenta fino a
quaranta volte. Per quanto concerne il controllo della temperatura, Mammiferi
ed Uccelli riescono a mantenere una temperatura costante, generalmente più alta
rispetto a quella esterna, e per questo si dicono Omeotermi. Fino
a pochi anni fa si pensava che i Rettili avessero una temperatura corporea
fluttuante in relazione a quella dell’ambiente, ed erano pertanto definiti Eterotermi,
ma attualmente si riconosce ai Rettili un certo controllo della temperatura.
Molto più importante è distinguere i Rettili da Mammiferi ed Uccelli sulla base
dell’origine della temperatura corporea. I Rettili mantengono la loro
temperatura corporea più o meno costante e al di sopra della temperatura
dell’ambiente assorbendo dall’esterno il calore delle radiazioni solari e, in
misura minore, ricavando calore dal loro metabolismo basale. Per questo motivo,
i Rettili sono definiti Ectotermi. Mammiferi ed Uccelli, invece,
mantengono la loro temperatura corporea costante e più elevata rispetto a
quella dell’ambiente, soprattutto grazie al calore prodotto dal loro
metabolismo basale, quindi vengono definiti Endotermi. Il grado
di attività mostrato dai Dinosauri pare essere più alto da quello dei Rettili
attuali: può forse essere correlata ad un metabolismo più elevato?
Per dare una risposta a
questa domanda, dobbiamo analizzare alcuni elementi.
1 –
Tutti i Dinosauri hanno
una postura più eretta rispetto ai Rettili attuali, che portano omero e femore
orizzontalmente rispetto al corpo. In questo carattere i Dinosauri sono più
vicini ai Mammiferi che non ai Rettili. Molti Dinosauri, inoltre, avevano la
possibilità di una stazione eretta, usando la coda come bilanciere, cosa che
non accade mai negli altri Rettili, che sono tutti quadrupedi. La camminata
quadrupede è messa in relazione, per i Rettili, ad un basso metabolismo.
Infatti, i Rettili attuali utilizzano per i movimenti un metabolismo
fermentativo, in cui l’attività muscolare prolungata porta ad un rapido
accumulo di acido lattico, fatto che porta all’inevitabile e frequente
sospensione dell’attività muscolare stessa per consentire il riassorbimento
dell’acido. Nessun rettile attuale può essere considerato un buon camminatore o
corridore, in contrasto con i Mammiferi, che presentano un metabolismo
ossidativo. I Dinosauri, per l’architettura scheletrica e per le tracce
fossilizzate lasciate sul terreno, si presentano come ottimi camminatori e
talvolta anche corridori. Periodi prolungati di attività muscolare avrebbero
richiesto uno spostamento verso un metabolismo ossidativo, con conseguenti
modificazioni al sistema digestivo, a quello respiratorio e a quello
circolatorio, ma
2 – ISTOLOGIA DELLE OSSA
Le ossa dei
Rettili attuali mostrano pochi canali haversiani ed un basso tasso di
rimodellamento: è pertanto possibile riconoscere delle “linee di accrescimento”
corrispondenti a momenti di temperatura corporea più o meno elevata.
Nelle ossa dei Mammiferi,
e in misura minore nei Dinosauri, non si notano linee di accrescimento, ma si
osserva un’alta densità di canali haversiani. Ciò suggerisce che i Dinosauri
avessero un metabolismo più elevato, un rimodellamento dell’osso più attivo,
temperatura corporea più alta ed uno scambio di minerali tra sangue ed ossa più
rapido ed efficiente. Conseguentemente, nulla vieterebbe di pensare, per i
Dinosauri, ad una temperatura corporea e ad un metabolismo più elevati rispetto
agli attuali Rettili. Rimane aperto il problema se l’istologia delle ossa sia
collegata ad una temperatura corporea più elevata, che potrebbe avere qualsiasi
origine, o ad un tasso metabolico maggiore, il che ci porterebbe verso
l’endotermia. Occorre comunque fare una distinzione all’interno dei Dinosauri
per quanto concerne l’istologia delle ossa: i grandi Sauropodi sono quelli che
più si avvicinano ai Mammiferi, mentre i Carnosauri e i piccoli Coelurosauri
sono più vicini ai Rettili. Gli attuali coccodrilli, unici rappresentanti
viventi della classe degli Arcosauri a cui appartenevano i Dinosauri, pur
essendo ectotermici, hanno caratteri istologici che si avvicinano a quelli dei
Mammiferi. L’analisi dell’istologia delle ossa dei Dinosauri ci porta a
concludere quindi che i grandi Sauropodi avevano una temperatura corporea più
alta e costante rispetto a Carnosauri e Coelurosauri, ma non ci dà precise
indicazioni di ectotermia o endotermia.
3 – STRUTTURA DELLA
COMUNITÀ
Se i Dinosauri,
come i Mammiferi endotermi, necessitano di almeno dieci volte la quantità di
cibo consumata dagli ectotermi, questo dovrebbe essere evidenziato dal rapporto
predatori/predatori+prede, espresso come biomassa.
Nelle comunità
Paleozoiche di Anfibi e Rettili ectotermici, questo rapporto è,
approssimativamente di uno a uno. Nelle comunità di Mammiferi attuali, i
predatori rappresentano circa il 3-5%
delle possibili prede. Nelle comunità di Dinosauri, questo rapporto si
avvicina a quello dei Mammiferi (5-10%), il che potrebbe essere un’indicazione
di endotermia per i Dinosauri, almeno per quelli carnivori. Bisogna però
ricordare che studi recenti condotti sul grande varano dell’isola di Komodo,
che è una forma ectotermica, hanno mostrato un rapporto predatori/prede simile
a quello dei Mammiferi.
4 – DIMENSIONI
Alcuni autori recentemente hanno suggerito che probabilmente i Dinosauri non avevano un metabolismo intermedio tra quello dei Rettili e quello dei Mammiferi, ma che la loro soluzione era del tutto originale. Ciò che più colpisce dei Dinosauri sono le dimensioni, talvolta veramente gigantesche: solo pochissime forme pesavano meno di cinque kg, mentre molte pesavano più di una tonnellata. Basti pensare che i grandi Sauropodi avevano un peso variabile tra le dieci e le 100 tonnellate. Gli attuali Mammiferi ed Uccelli si caratterizzano, al contrario, per le loro dimensioni abbastanza contenute. L’effetto più importante delle grandi dimensioni risiede nel rapporto fra perdita e acquisto di calore con l’ambiente. A causa dell’elevato rapporto superficie/volume, i piccoli Rettili attuali perdono rapidamente calore corporeo durante la notte e i periodi freddi, e non sono pertanto capaci di uno sforzo prolungato. Al contrario, le forme rettiliane di grandi dimensioni (tartarughe, coccodrilli, varani), grazie al favorevole rapporto tra superficie e volume del corpo, presentano oscillazioni della temperatura corporea più contenute. Animali di dimensioni enormi, quali erano i Dinosauri, potrebbero avere avuto un ottimo controllo della temperatura corporea senza avere bisogno di un metabolismo troppo elevato e paragonabile a quello dei Mammiferi, sfruttando la cosiddetta Omeotermia inerziale. Animali delle dimensioni di un sauropode con un livello metabolico paragonabile a quello di un mammifero, avrebbero avuto necessità di particolari strutture, come ghiandole sudoripare, per disperdere il calore, ma queste non traspaiono dai resti fossili (abbiamo anche l’impronta della pelle dei Dinosauri). La temperatura costante raggiunta attraverso l’aumento delle dimensioni avrebbe offerto i vantaggi dell’omeotermia dei Mammiferi, ma ad un costo, in termini di consumo di cibo, molto inferiore.
Vantaggi nei Mammiferi, l’80-90% del cibo consumato è utilizzato, dalle forme di piccole dimensioni, per mantenere una temperatura corporea abbastanza alta e costante. L’endotermia permette ai Mammiferi un’ampia distribuzione geografica. Questi vantaggi sono bilanciati dal fatto che essi debbono consumare dieci volte più cibo di forme ectotermiche dello stesso peso.
Si può pensare che il gigantismo abbia rappresentato la “scappatoia” più semplice per mantenere una temperatura corporea costante e più alta rispetto all’ambiente, e ciò ad un costo metabolico basso. Questa scelta rende però l’organismo estremamente vulnerabile ad ogni deterioramento climatico. Così, quando le variazioni di temperatura, tanto in positivo quanto in negativo, avessero superato una certa soglia per un periodo abbastanza prolungato, avrebbero rappresentato un fattore “fatale” per l’individuo.
Gli esemplari delle poche specie di piccole dimensioni e gli esemplari giovani non sarebbero logicamente stati omeotermi inerziali, ma sarebbero stati soggetti ad una rapida perdita di calore e pertanto avrebbero dovuto regolare la temperatura corporea utilizzando le risorse ambientali.
Rimane evidente che la soluzione qui prospettata per la fisiologia dei Dinosauri non può che ritenersi, anche se logica, puramente speculativa. Tra gli attuali Vertebrati esiste una chiara distinzione tra eterotermi ed omeotermi, ma attualmente non abbiamo alcuna possibilità di studiare forme che presentino omeotermia inerziale.
CONSIDERAZIONI
FINALI
Secondo la prima ipotesi, cioè quella prospettata da Alvarez, si avrebbe avuto un improvviso raffreddamento del pianeta, che sarebbe durato solo alcuni mesi, che avrebbe causato una drastica riduzione della vegetazione. Questo periodo sarebbe stato seguito da un altrettanto notevole innalzamento della temperatura a causa dell’aumento dell’anidride carbonica. L’effetto del raffreddamento avrebbe dovuto farsi sentire su ogni essere vivente, non soltanto su quelli di maggiori dimensioni, quali Saurischi ed Ornitischi, e su alcune forme medio-piccole, quali i Marsupiali. Inoltre, se il famigerato meteorite fosse stato l’unico responsabile dell’estinzione, questa avrebbe dovuto essere improvvisa e non diluita nel tempo. Oggi noi sappiamo con certezza che sulla terraferma si estinsero, tra i Rettili, i Saurischi, gli Ornitischi e gli Pterosauri, mentre altre forme (coccodrilli, tartarughe e lacertidi) non subirono alcuna crisi. Inoltre, da alcuni recenti studi, si evince che il fenomeno finale non sia stato improvviso, ma si sia protratto per almeno duecento mila anni, inducendo a pensare che l’estinzione sia stata dovuta ad un graduale cambiamento climatico. Ed i Dinosauri, con le loro grandi dimensioni e la loro omeotermia inerziale, risultano i più esposti ai cambiamenti climatici, sia in positivo che in negativo. Gli altri rettili di piccole e medie dimensioni hanno ancora oggi nel meccanismo di ibernazione/estivazione una naturale difesa contro le variazioni climatiche (un dinosauro non avrebbe mai potuto trovare o scavarsi una tana abbastanza grande in cui svernare). Anche tra i Mammiferi l’estinzione fu, in un certo senso, selettiva: fu più pesante tra le forme con endotermia meno perfezionata (Multitubercolati e Marsupiali), e più leggera per quelle già più spiccatamente endotermiche (Placentati).
A cosa bisogna imputare quindi l’estinzione di massa di fine Mesozoico?
La grande estensione dei mari durante tutto il Cretaceo aveva instaurato su tutti i continenti un clima fortemente influenzato dalle grandi masse acquee, con scarsa stagionalità e temperature medie più alte di quelle attuali. Verso la fine del Cretaceo, si verificò un aumento dell’attività tettonica, che portò variazioni nella circolazione delle correnti marine. Conseguenza principale della caduta del livello marino fu la continentalizzazione del clima, mentre il cambiamento di circolazione delle acque portò ad un abbassamento della temperatura media stimato, negli Stati Uniti, dell’ordine di dieci gradi centigradi. Questo abbassamento della temperatura e l’instaurarsi di una stagionalità con inverni freddi ed estati calde è sufficiente per spiegare la grande rivoluzione che si nota nelle flora, ma soprattutto nella fauna e che abbiamo appena descritto. Non sembra pertanto necessario invocare anche un fattore extra terrestre per spiegare la crisi biologica della fine del Cretaceo, in quanto l’interpretazione del succedersi degli eventi è sufficiente a darne una motivazione plausibile.
Sia comunque chiaro che non si intende in questa sede sottovalutare la presenza dell’Iridio al limite K/T: esso è il testimone di un evento catastrofico contemporaneo al culmine della crisi biologica; di questo evento catastrofico è comunque difficile valutare gli effetti su una fauna la cui biologia è soltanto ricostruibile per supposizioni e non basandosi su correlazioni con le faune attuali.
DINOCURIOSITÀ
IL
PIÙ LUNGO
Seismosauro:
IL PIÙ PICCOLO
Compsognato:
IL PIÙ VELOCE
Gallimimo e ornitomimo:
IL
CARNIVORO PIÙ GRANDE
Gigantosauro:
IL
PIÙ LONGEVO
Tutti
i Sauropodi, circa 100 anni
IL
COLLO PIÙ LUNGO
Mamanchisauro:
Diplodoco:
IL
MAGGIOR NUMERO DI DENTI
Gli
Adrosauri, con circa 96 denti
IL
PIÙ LENTO A «DIVENTARE GRANDE»
Botriospondilo: impiegava 43 anni per raggiungere le dimensioni dell’adulto
IL
CRANIO PIÙ GRANDE
Triceratopo:
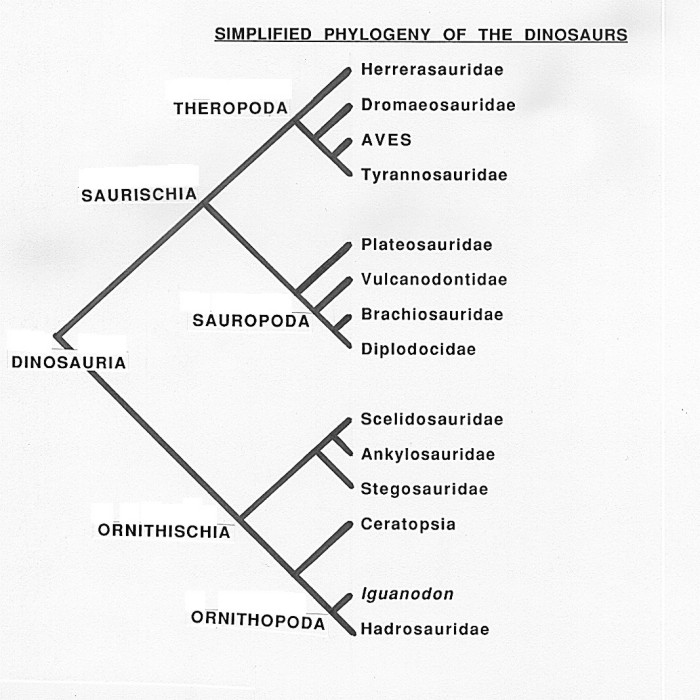
Albero filogenetico semplificato dei Dinosauri
BIBLIOGRAFIA
Letture consigliate di Paleontologia generale:
Beneš, Josef, 1982 – Animali e piante della
preistoria, Fabbri Editori, Milano.
Colbert, Edwin H., 1973 – Animali e continenti alla
deriva, Arnoldo Mondadori Editore, Milano.
de Groot, G. E., 1972 – I fossili raccontano (titolo
originale: Levensvormen uit het verleden), Zanichelli, Bologna.
Kirkaldy, J. F., 1978 – Fossili a colori, Edizioni
Paoline, Roma.
Moore, Ruth, 1954 – Uomo, tempo e fossili storia
dell’evoluzione, Garzanti, Milano.
Paul, Chris, 1982 – Storia naturale dei fossili,
Etas libri.
Pinna, Giovanni, 1974 – Alla ricerca dei fossili,
Longanesi, Milano.
Raffi, Sergio &
Serpagli, Enrico, 1993 – Introduzione alla Paleontologia, collana Scienze della Terra,
UTET, Torino.
Schneipp, Hermann, 1978 – I fossili, Editrice
Sirotti, Achille – appunti di Paleontologia dei
Vertebrati, Università degli Studi di Modena.
Špinar, Zdenĕk V.,
1972 – Quando
l’uomo non c’era (illustrazioni di Burian,
Zdenĕk), Fratelli Fabbri Editori, Milano.
Zorzin, Roberto, 2001 – Fossili per conoscere il
nostro passato, collana Piccole perle, Demetra, Verona.
Letture consigliate sull’argomento:
Aa. Vv., 1977 – L’avventura dei dinosauri, Arnoldo Mondadori
Editore, Milano.
Aa. Vv., 2001 – Dinosauri, numero
speciale della rivista “Newton”, RCS, Milano.
Benton, Mike, 1999 – Nel mondo dei dinosauri, i
fatti, BBC Worldwide Limited.
British Museum (a cura del), 1982 – I
Dinosauri e i loro parenti viventi, Cambridge University Press.
Haines, Tim, 1999 – Nel mondo dei Dinosauri –
immagini dalla preistoria, DeAgostini, Milano.
Lambert, D., 1991 – I Dinosauri dall’A alla Z,
realtà e leggenda delle più grandi creature del mondo preistorico, A. Vallardi.
Parker, Steeve, 1988 – Dinosauri dove e come
vivevano, Arnoldo Mondadori Editore, Milano.
Rigutti, Adriana, 1993 – Dinosauri, Atlanti
Scientifici Giunti.
Letture consigliate sull’estinzione dei Dinosauri:
Alvarez,
2001 – T. Rex e il cratere dell’apocalisse, Arnoldo
Mondadori Editore (titolo originale: T. Rex and the crater of
doom), Milano.
Hsü, Kenneth J., 1993 – La grande moria dei
Dinosauri (titolo originale: The Great Dying), Adelphi.
Michael,
Jean-Guy, 1993 – I Dinosauri misteri di una scomparsa (titolo
originale: Le mond perdu des dinosaures),
Electa/Gallimard.
Letture varie e
di approfondimento:
Carlini,
F., 1993 – Il ritorno dei DNAsauri, i segreti di Jurassic
Park, Manifesto libri.
Kent,
George C., 1997 – Anatomia comparata dei Vertebrati, Piccin, Padova.
Su Internet:
http://Sandia.gov/media/dinosaur.htm
http://www.mediasoft.it/dinosuri/pages/p1.htm
http://members.it.tripod.de/LORY
88/dino.htm
http://www.areacom.it/arte_cultura/dinosauri
http://spazioitalia.com/fossili/index.htm
http://www.hcc.hawaii.edu/dinos/dinos1.html
http://www.babyonweb.com/giochi/dinosauri.htm
http://www.nationalgeographic.com/dinorama
http://www.dinosaur.org/frontpage.html
http://www.search4dinosaurs.com/pictures.html
http://www.tarantasio.it/pangea/homepage.asp
http://www.arts-letters.com/dino2/ency/index.html
http://www.ucmp.berkeley.edu/diapsids/dinolinks.html#art
http://www.EnchantedLearning.com/subjects/dinosaurs/index.html
Ringraziamenti finali
Desidero
ringraziare infinitamente il prof. Maurizio Gnoli, che mi ha inculcato l’amore
per
|
Milioni
di anni |
Era |
Periodo |
Epoca
|
|
|
23,5 |
Terziario |
Paleogene |
Oligocene |
|
|
35 |
Eocene |
|||
|
65 |
Paleocene |
|||
|
88 |
Mesozoico |
Cretaceo |
Senoniano |
|
|
130 |
Gallico |
|||
|
145 |
Neocomiano |
|||
|
157 |
Giurassico |
Malm |
||
|
180 |
Dogger |
|||
|
210 |
Lias |
|||
|
235 |
Triassico |
Tr
3 |
||
|
243 |
Tr
2 |
|||
|
245 |
Scitiano |
|||
|
Paleozoico |
||||