| CSEI 2002 |
|
|
| Corso "Metodi avanzati nella potabilizzazione delle acque" Taormina 4-6 Novembre 2002 |
|---|
|
Microrganismi patogeni emergenti veicolati dall'acqua e nuovi metodi molecolari per il controllo della contaminazione microbica delle acque G.M. Giammanco |
| Dipartimento di Igiene e Microbiologia "G. D'Alessandro", Università degli Studi di Palermo, via del Vespro 133, 90127 Palermo |
|---|
| L'accesso all'acqua pulita costituisce un bisogno umano primario,
quindi un diritto fondamentale dell'uomo. L'acqua contaminata rappresenta un serio
pericolo per la salute sia fisica che sociale dellíintera popolazione. L'assenza
d'acqua sicura è un'offesa per la dignità umana. Kofi Annan (Segretario Generale O.N.U.) |
|
|
|
| Paesi in via di sviluppo: 1) Malattie trasmesse con l'acqua (contaminazione) 2) Malattie dovute a carenza d'acqua (scarsa igiene) 3) Malattie ospitate dall'acqua, l'acqua costituisce l'habitat per organismi nei quali alcuni parassiti trascorrono parte del loro ciclo vitale. 4) Malattie collegate all'acqua, l'acqua fornisce l'habitat per insetti vettori. Paesi sviluppati: 5) Infezioni propagate dall'acqua, microrganismi che proliferano nelle condutture dellíacqua potabile e possono contaminare l'organismo attraverso le vie respiratorie (legionella). |
|
Nuovi patogeni trasmissibili con l'acqua riconosciuti negli ultimi 25 anni:
|
| Caratteristiche dei nuovi patogeni emergenti: Negli individui adulti in buona salute: Determinano malattie raramente debilitanti per non più di poche ore o qualche giorno Nei bambini più piccoli, negli anziani e negli individui immunocompromessi o debilitati: Possono causare malattie severe e anche la morte Sono difficili da individuare con le classiche metodiche microbiologiche In alcuni casi mostrano una certa resistenza o refrattarietà ai comuni metodi di sanificazione delle acque (cloro) |
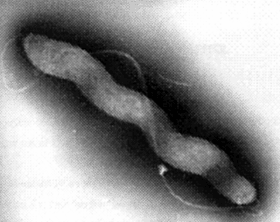
Campylobacter jejuni
|
Negli ultimi anni:Prima causa di gastroenterite nei paesi sviluppati. Serbatoio dell'infezione:
Frequenza di isolamento dalle acque superficiali: |
|
Modalità di infezione:
Periodo di incubazione: 1-10 gg. L'eliminazione del microrganismo con le feci dura fino a 8 settimane dallíinizio dei sintomi. Dopo l'infezione si sviluppa una immunità duratura. Tuttavia,
la presenza di diversi sierotipi consente il verificarsi di infezioni ripetute nello
stesso soggetto |
Cryptosporidium parvum
|
|
|
|
Cryptosporidium (a) la prevalenza di Cryptosporidium nell'acqua di fonte è alta; (b) le oocisti di Cryptosporidium sono refrattarie al trattamento con cloro dellíacqua potabile; (c) i metodi di filtrazione utilizzati normalmente per le acque potabili di superficie non rimuovono efficientemente le oocisti di Cryptosporidium, a causa del loro piccolo diametro; (d) la carica infettante di Cryptosporidium per gli esseri umani è estremamente bassa. Anche se carenze nel trattamento delle acque sono state identificate
in alcune epidemie, almeno un'epidemia si è presentata in rapporto con una
moderna stazione di trattamento delle acque, in cui il trattamento era ben documentato
ed effettuato in maniera corretta. Negli Stati Uniti, il controllo sistematico della
presenza di Cryptosporidium nell'acqua potabile è obbligatorio per
tutti gli impianti di trattamento e distribuzione che servono una popolazione superiore
alle 100.000 persone. Tuttavia, i metodi correnti utilizzati per la ricerca di Cryptosporidium
possono sottostimare la presenza del parassita. In avvenire, metodi di analisi più
sensibili basati sulla ricerca del DNA del parassita possono facilitare la rilevazione
di Cryptosporidium nei campioni ambientali.
|
 |
|
Legionella
Questi batteri causano una malattia respiratoria nell'uomo quando
un ospite suscettibile inala aerosol o aspira acqua contaminata. La malattia che
ne deriva, la legionellosi, classicamente si presenta come due entità cliniche
distinte, la malattia dei Legionari, una malattia severa multisistemica che comprende
la polmonite, e la febbre di Pontiac, una malattia auto-limitante simil-influenzale.
Inoltre, in molte persone líinfezione avviene in forma assolutamente asintomatica.
Non è possibile distinguere clinicamente la malattia dei Legionari da altri
tipi di polmoniti, anche se all'esame radiografico del torace gli infiltrati alveolari
sono più comuni con la malattia di Legionari. La certezza diagnostica si ottiene
solo con la dimostrazione della presenza del microrganismo nei campioni clinici. Questi includono: (a) la presenza del batterio in un ambiente acquatico, (b) la possibilità per il batterio di moltiplicarsi fino a raggiungere la carica infettante necessaria, (c) la trasmissione dei batteri via aerosol ad un ospite umano
suscettibile all'infezione. Colonie di L. pneumophila su charcoal yeast
extract agar L'esame microscopico dei campioni dopo colorazione con anticorpi fluorescenti specifici (direct fluorescent antibody = DFA) è stato il primo metodo impiegato per rilevare la presenza di legionelle nel tessuto polmonare e nelle secrezioni respiratorie. Il metodo DFA inoltre può essere usato per la tipizzazione sierologica degli isolati di Legionella. Un numero limitato di sieri policlonali coniugati con fluorescina è disponibile per riconoscere i sierogruppi di L. pneumophila e le specie di L. non-pneumophila. La specificità e la sensibilità di questi reagenti non sono state ancora ben valutate, ma sono state segnalate reazioni crociate con batteri non-Legionella.
Ricerca dell'acido nucleico di Legionella. La prima analisi progettata per rilevare la presenza di DNA di L. pneumophila era basata sullíutilizzo di una sonda radioattiva specifica per il DNA ribosomico, comune a tutte le specie di Legionella. Studi successivi hanno segnalato la scarsa specificità di questa analisi ed il kit diagnostico che la impiegava è stato ritirato dal mercato. La PCR rappresenta uno dei pochi test diagnostici in grado di rilevare le infezioni causate da tutte le specie conosciute di Legionella. Tuttavia, la maggior parte dei kit in commercio rilevano soltanto le infezioni dovute al sierogruppo 1 di L. pneumophila. I diversi kit di PCR che sono stati sviluppati per legionella utilizzano come sequenze bersaglio, il gene per lírRNA 5S, il gene per lírRNA 16S, o il gene di mip. Uno dei kit più ampiamente usati per rilevare la presenza di legionelle nell'ambiente (kit EnviroAmp; Perkin-Elmer, Inc., Norwalk, Connett.), amplifica simultaneamente due bersagli, un bersaglio nellírRNA 5S specifico per il genere di Legionella e una parte del gene di mip specifica per L.pneumophila. tuttavia anche questo kit è stato ritirato dal mercato nel 1997 a causa della sua scarsa sensibilità, nonostante un'alta specificità. Altre analisi in PCR che utilizzano come gene bersaglio lírDNA 16S si sono dimostrate altamente sensibili e specifiche. Tuttavia, un certo numero di risultati falso-positivi sono stati ottenuti con specie di Acinetobacter e con un Proteobacterium non identificato e con specie appartenenti al genere Gemella. |
|
METODI DIAGNOSTICI INNOVATIVI PER LA RICERCA DI MICRORGANISMI PATOGENI NELLE ACQUE |
|
Figura 5. Separazione dei prodotti amplificati da un campione
misto usando la RP-IP HPLC. I singoli ampliconi vengono isolati fisicamente dalla
mistura utilizzando un collettore per recuperare dalla colonna líeluente che corrisponde
a ogni picco rilevato.
Figura 7. Electroferogrammi della regione ipervariabile
dellíamplicone ottenuto da un isolato ambientale. I prodotti di PCR vengono tagliati
con una serie di endonucleasi di restrizione ed i frammenti risultanti vengono separati
con un sistema di elettroforesi capillare in base alla taglia. La presenza e la taglia
di ogni frammento di restrizione forma il "fingerprint" pattern dal quale
si deduce líidentità dellíisolato per confronto con una banca dati di pattern
di restrizione.
Figura 8. Cromatogramma di una sequenza parziale di un ceppo
di E. coli. Il sequenziamento fornisce molte più informazioni rispetto
all'analisi dei pattern di restrizione. Esso può essere utilizzato anche per
identificare nuovi microrganismi, che non sono compresi nelle banche dati di pattern
di restrizione.
Figura 12. Popolazione batterica mista di Comamonas acidovorans ed Escherichia
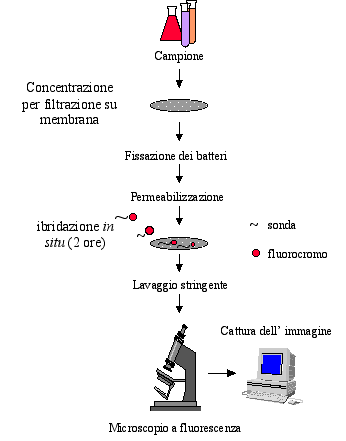
coli (ratio 1 : 4000) evidenziata con sonde specifiche. BIBLIOGRAFIA |
| |
Go to the Home Page |