

Le micorrize
Sono associazioni simbiontiche che si instaurano tra radici di molte piante e funghi
del sottosuolo (dal greco: mykos: fungo e rhiza: radice).
Benché se ne senta parlare troppo poco, è il tipo di simbiosi più diffuso in natura:
più del 90% delle specie vegetali in condizioni naturali risulta micorrizato.
Addirittura, sono stati trovati resti fossili che confermano l’esistenza delle
endomicorrize già 450 milioni di anni fa, contemporaneamente all’apparizione dei
vegetali sulle terre emerse. Si ritiene che siano state fondamentali nel processo di
colonizzazione dei continenti, e a tutt’oggi sono ancora necessarie per la
vegetazione contemporanea. Tuttavia negli ambienti antropizzati (campi coltivati e verde
urbano) le micorrize sono spesso assenti oppure presenti in forma molto ridotta, molto
probabilmente a causa dell’inquinamento chimico dei terreni.
L'esistenza della simbiosi micorrizica fu notata per la prima volta nel 1883 da Gibelli
presso L'Orto Botanico di Torino, ma fu B. Frank che nel 1885 definì micorriza questa
struttura. I caratteri di queste micorrize così come li ha individuati Frank consistono
di una sorta di rivestimento, o mantello, costituito da un intreccio ifale, che include o
riveste le ultime terminazioni radicali. Frank notò che queste strutture sono sempre
presenti nei sistemi radicali delle piante in ambienti naturali. Successivamente furono
individuati due tipi di micorrize: le ECTOMICORRIZE,
caratteristiche della maggior parte delle latifoglie e delle conifere, dotate di un
mantello fungino esterno ricoprente l’apice radicale, e le
ENDOMICORRIZE, a più ampia diffusione (anche tra le specie erbacee), non
dotate di un mantello fungino esterno, ma presenti anche all’interno delle cellule.
Le ectomicorrize interessano un numero relativamente limitato di specie vegetali (per lo
più piante arboree forestali) ed un elevato numero di specie fungine.
Al contrario, le endomicorrize interessano il 90% dei vegetali, ed un numero limitato di
specie fungine (classe Zygomicetes).
ENDOMICORRIZE |
ECTOMICORRIZE |
SEZIONE
DI APICE RADICALE |
ECTOMICORRIZE
Sono le micorrize tipiche dei tartufi e dei porcini, comparse sulla terra probabilmente
intorno a 150 milioni di anni fa.
Sono così definite perché il fungo non penetra mai all’interno delle cellule
dell’ospite. Le ife fungine formano come uno spesso strato attorno alle radici, detto
mantello o micoclena. Colore, spessore, morfologia del mantello possono variare a seconda
delle specie Dal mantello le ife si insinuano tra le cellule della corteccia radicale,
formando un intreccio intercellulare, il reticolo di Hartig. A seconda dell’ospite
questo reticolo può essere più o meno sviluppato, e raggiungere il cilindro centrale
(conifere) oppure limitarsi ai primi strati cellulari della corteccia (latifoglie). Sempre
dal mantello si diparte una fitta rete di ife esterne e cordoni miceliari che si estendono
notevolmente nel suolo circostante, e in condizioni appropriate possono produrre strutture
riproduttive (carpofori: es. tartufi e funghi)
Generalmente la radice micorrizata risulta profondamente trasformata: la crescita è
bloccata e si biforca, e assume una forma coralloide. Il mantello fungino spinto dalle
radichette secondarie non si rompe, ma si estende entrando in attiva proliferazione
cellulare, inglobando le nuove radici laterali. Nella radice micorrizata scompaiono i peli
radicali.
Queste micorrize sono essenzialmente formate da circa 4–5.000 specie fungine
appartenenti agli Ascomiceti e Basidiomiceti, e sono presenti nelle piante forestali
(latifoglie e conifere).
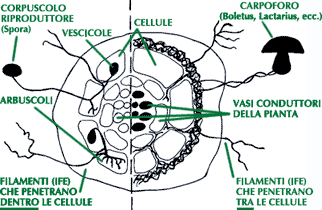
ENDOMICORRIZE ARBUSCOLARI
Sono così definite perché, a differenza delle ecto, il fungo penetra all’interno
delle cellule dell’ospite, mentre mancano di un mantello fungino esterno (si parla di
funghi VAM). La comparsa di queste micorrize sulla Terra si suppone risalga almeno a 400
milioni di anni fa, sono perciò più antiche e più affermate delle ectomicorrize.
Le spore che si trovano nel terreno germinano in presenza di radici ospiti per effetto
degli essudati radicali. Si sviluppano fino a raggiungere la radice stessa, e la
colonizzano penetrando sia attraverso gli spazi intercellulari sia direttamente nelle
cellule. Il fungo si diffonde così attraverso le cellule corticali, senza invadere mai il
cilindro centrale e le cellule dell’apice radicale.
All’interno delle cellule le ife si diramano a formare delle strutture ramificate,
gli arbuscoli, responsabili degli scambi nutrizionali tra i due simbionti: la pianta cede
i carboidrati eccedenti prodotti attraverso la fotosintesi, il fungo a sua volta cede i
sali minerali assorbiti dal suolo circostante. Gli arbuscoli hanno vita breve: dopo pochi
giorni infatti degenerano. Un’altra struttura prodotta dalle ife fungine è la
vescicola, rigonfiamento tondeggiante inter o intracellulare, organo di accumulo di
granuli di grasso con funzione di riserva.
Le spore formate dai funghi AM sono asessuali, e si formano dalle ife vegetative.
Parecchi studi dimostrano che, seppur la radice non subisce variazioni morfologiche
notevoli come avviene per le ectomicorrize, l’apparato radicale risente della
presenza del fungo AM: possono infatti variare il grado di ramificazione e le dimensioni
delle radici stesse, fino ad aumentare d centinaia di volte.
Formano endomicorrize arbuscolari circa 130 specie di funghi appartenenti all’ordine
delle Glomales (Zygomiceti), e interessano molte piante erbacee, ortive, da frutto,
forestali e tropicali, indice di mancanza di specificità. Si trovano anche presso le
Pteridofite e Epatiche. In ambiente asfittico non esistono.
Esistono altri tipi di micorrize: le micorrize delle ericacee, le ectoendomicorrize
(arbutoidi o monotropoidi), le endomicorrize delle orchidee (si tratta di
particolari micorrize endotrofiche, essenziali anche per la germinazione del seme che può
avvenire solo in presenza di una infestazione di ife fungine).
FISIOLOGIA DELLE MICORRIZE
Non solo la morfologia della pianta ospite viene modificata dall’instaurarsi della
micorriza, ma anche la fisiologia risulta profondamente alterata.
Brevemente, la pianta ospite cede al fungo zuccheri e vitamine, mentre il fungo assorbe e
trasferisce alla pianta gli elementi minerali. Infatti lo sviluppo considerevole delle ife
extramatricali nel terreno permette di esplorare un volume di suolo notevolmente maggiore
di quanto può fare la singola radice, anche lontano dalla zona di assorbimento della
radice stessa, aumentando notevolmente la quantità di sostanze nutritive raggiungibili.
Non solo, ma le micorrize sono in grado di solubilizzare e quindi assorbire le forme
organiche o minerali presenti nel suolo in composti insolubili, non direttamente
utilizzabili dalle piante, cambiando quindi radicalmente il conto agronomico della
disponibilità degli elementi nutritivi nel terreno.
Il maggior assorbimento di sali minerali dal suolo (P, N, Ca, K, Fe, Mg, Cl, Zn, Cu)
modifica l’equilibrio e la composizione dei nutrienti nei tessuti della pianta con
conseguenze sulla resa fotosintetica che viene aumentata e sulla ripartizione del
fotosintetato tra radice e parte aerea della pianta. Il 20% del C totale assimilato dalla
pianta può essere trasferito al fungo.
Tutti questi mutamenti di assorbimento e distribuzione dei nutrienti portano ad un
cambiamento dell’essudazione radicale, e l’aumentato flusso di C verso il suolo
mediato dalle micorrize è importante per l’ecosistema, e può essere considerato un
altro dei benefici dovuto alle micorrize. Anche il ritorno di sostanza organica al suolo
viene incrementato con la micorrizazione, accompagnato anche da un maggior rilascio di N,
P e K.

![]()