SCUOLA DI SPECIALIZZAZIONE IN MICROBIOLOGIA E VIROLOGIA
Direttore Prof. Alfredo Chiarini
IDENTIFICAZIONE DI CANDIDA DUBLINIENSIS IN UN GRUPPO DI PAZIENTI HIV +
Tesi di specializzazione di: Sonia Pecorella
INTRODUZIONE
1. Le micosi
opportunistiche.
Il Regno dei Funghi è incredibilmente eterogeneo e i suoi membri, organismi unicellulari, eucarioti, aerobi o anaerobi facoltativi, occupano uno straordinario numero di nicchie ecologiche, vivendo negli strati superficiali del suolo o come commensali di vari organismi vegetali ed animali. Nonostante questa loro notevole versatilità, le specie che nell’uomo riescono a causare malattie (le cosiddette micosi) sono relativamente poche. Alcune di esse sono considerate dei patogeni opportunisti.
Con il termine opportunismo si indica il comportamento di quei microrganismi che, pur essendo abitualmente incapaci di provocare uno stato morboso, possono tuttavia causare un’infezione quando sia loro offerta una “opportunità”, fondamentalmente rappresentata da una riduzione delle capacità difensive dell’organismo ospite.
Le infezioni che riconoscono questa particolare patogenesi prendono il nome di infezioni opportunistiche.
Negli ultimi anni, la loro frequenza è aumentata drammaticamente in patologia umana, creando rilevanti problemi sia di ordine clinico che epidemiologico e trovando ampia espressione nel campo delle micosi.
I principali miceti opportunisti sono elencati nella tabella 1.1. [1]. Le specie segnalate più di frequente appartengono ai generi Candida e Aspergillus [2]. Le malattie prodotte da questi microrganismi possono interessare l’apparato respiratorio (Aspergillosi) o le mucose e la cute (Candidosi).
Tabella
1.1. I miceti opportunisti.
__ · Candida
· Cryptococcus
________________________________ Dati modificati da Dean e Burchard.1 |
2. Il genere Candida.
Candida è un fungo lievitiforme, dotato quindi di tallo (corpo del micete) unicellulare, costituito da cellule isolate rotondeggianti od ovali (blastospore). A causa della scarsa coerenza delle cellule che lo costituiscono, le colonie di questo micete hanno aspetto e consistenza cremosa o pastosa, che ricorda quella abituale delle colonie batteriche.
La riproduzione avviene per gemmazione: si forma nuovo materiale cellulare, aumentano le dimensioni della cellula e da una protuberanza del tallo fungino, in seguito alla formazione di un setto di divisione, si originano due blastospore. Queste ultime possono andare incontro a gemmazione successiva, allungandosi e rimanendo unite tra loro a formare pseudoife (eccezionalmente anche ife vere). Inoltre, è possibile in alcune specie osservare la formazione di tubuli germinativi e clamidospore apicali sferiche, isolate o a grappoli.
Candida è un commensale della cute e delle mucose dei tratti gastrointestinali e urogenitali dell’uomo, ma è stato anche ritrovato nell’ambiente, in particolare sulle foglie, sui fiori, nell’acqua e nel suolo.
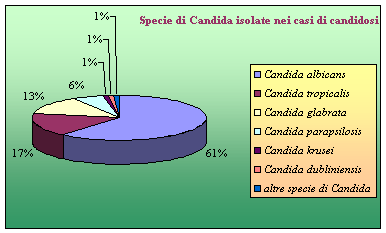
Si conoscono circa 200 specie appartenenti al genere Candida [3] ma solo il 10% di queste sono in grado di provocare malattie nell’uomo, purché coesistano delle condizioni predisponenti. Si tratta infatti di funghi opportunisti per eccellenza. Le specie isolate più di frequente nei casi di infezione e la loro incidenza sono indicate nella grafico 1.1.
|
Grafico 1.1. Specie patogene più comuni del
genere Candida (dati modificati da Dean e
Burchard1). |
Oltre a C. albicans, che è la specie isolata nella maggior parte dei casi di candidosi, una particolare patogenicità è ammessa per almeno altre cinque specie: C. tropicalis, C. glabrata, C. parapsilosis, C. krusei e C. dubliniensis, riunite sotto il termine “specie Candida non-albicans”.
Recentemente, si è assistito ad un incremento delle infezioni dovute alle specie appartenenti a questo gruppo [4,5].
2.1. La patogenesi.
Generalmente le infezioni causate da Candida spp. sono indicate con il termine di candidosi. I quadri clinici possono essere estremamente diversi, così come gli organi “bersaglio”.
La candidosi può essere superficiale e localizzata o profonda e disseminata. Nelle forme superficiali, quelle più comuni, le lesioni interessano le mucose (candidosi orale, vulvovaginiti), più di rado la cute (intertrigine) e le unghie (onicomicosi da Candida). Nelle forme disseminate invece, l’infezione può diffondersi dal primo focolaio infiammatorio ad altri organi e tessuti per via ematica o linfatica, dando luogo alle più svariate localizzazioni solitarie o multiple, con quadri clinici gravi o gravissimi (pneumopatie, pielonefriti, forme viscerali e sistemiche, etc.).
Il passaggio di Candida dalla situazione di commensalismo all’infezione è costantemente mediato da una serie di fattori favorenti, molti dei quali sono indicati nella Tabella 2.1.1.
FATTORI PREDISPONENTI
|
ESEMPI
|
Fisiologici |
Gravidanza,
età (anziani e bambini) |
Traumi |
Macerazioni,
infezioni, ustioni, piaghe |
Ematologici |
Neutropenia,
immunosoppressione cellulare (leucemia, linfoma, AIDS, anemia aplastica) |
Endocrini |
Diabete
mellito, ipoparatiroidismo, malattia di Addison |
Iatrogeni |
Chemioterapici,
corticosteroidi, contraccettivi orali, antibiotici, cateteri, protesi, interventi
chirurgici |
Altri |
Somministrazione
di droghe endovena, malnutrizione, malassorbimento |
Tabella 2.1.1. Fattori che predispongono
l’insorgenza della candidosi.
La candidosi è principalmente un’infezione endogena, quindi la sua origine è dovuta quasi sempre ad un’eccessiva crescita del micete in seguito ad una riduzione della reattività delle difese dell’individuo. Sono presenti, solitamente, uno o più fattori predisponenti rappresentati da traumatismi della cute o delle mucose, oppure da malattie preesistenti che determinano immunosoppressione (fattori ematologici: neoplasie maligne, AIDS) o iperglicemia (fattori endocrini: diabete) [6,7].
Anche alcune situazioni fisiologiche, come la gravidanza o le età estreme della vita, predispongono alla candidosi, la cui insorgenza è facilitata anche da fattori iatrogeni, quali alcune terapie farmacologiche (antibiotici a largo spettro, farmaci immunosopressori o corticosteroidi), interventi chirurgici (endolftalmiti susseguenti a trapianto di cornea da donatore infetto), presenza di cateteri e protesi, o somministrazione di droghe endovena [8,9,10]. Una candidosi orale si osserva spesso in bambini nati da madre con candidosi vaginale.
2.1.1. I determinanti di
patogenicità.
Soltanto negli ultimi anni sono stati messi in luce i meccanismi che determinano la virulenza di C. albicans e delle specie Candida non-albicans. La capacità di riconoscere ed aderire ai tessuti dell’ospite, di rispondere rapidamente ai cambiamenti ambientali e di secernere enzimi proteolitici e lipolitici sono oggi i fattori ritenuti maggiormente responsabili dell’azione patogena di questi miceti [11,12].
2.1.1.1. L’adesività.
Una fondamentale proprietà di Candida spp. è rappresentata dalla sua capacità di aderire specificamente e selettivamente alla superficie delle cellule di vari epiteli mucosi, grazie all’azione delle adesine [13,14]. Tale adesione viene considerata come la principale tappa del processo infettivo.
Notevole importanza sembrano rivestire anche le numerose interazioni specifiche di Candida con altri microrganismi o con la superficie delle protesi. Ad esempio, è stato dimostrato che C. dubliniensis è capace di coaggregare con Fusobacteium nucleatum, un batterio residente del cavo orale, mentre C. tropicalis può interagire con Streptococcus gordonii [15] e tali interazioni potrebbero facilitare la sopravvivenza di questi miceti nell’ambito della comunità microbica orale. In aggiunta, alcune specie di Candida, in particolar modo C. albicans, possono formare biofilm sulla superficie di protesi e di cateteri, riuscendo così a sfuggire all’azione antimicotica di alcuni farmaci (incremento della resistenza) [16-19].
2.1.1.2. La risposta ai cambiamenti
ambientali.
Sulla superficie delle mucose la limitazione dei nutrimenti, la competizione con le altre specie microbiche commensali e i meccanismi di difesa dell’ospite determinano una pressione selettiva che privilegia i microrganismi più adatti.
E’ noto che molti batteri sono in grado di regolare il proprio pathway metabolico in situazioni di stress, come ad esempio, durante la carenza di nutrienti. Sistemi di regolazione analoghi probabilmente esistono anche nel genere Candida ma non sono stati ancora caratterizzati. Si sa, infatti, che molti geni Candida-specifici codificano per delle proteine cataboliche e si può ipotizzare che alcuni di questi geni siano implicati direttamente nei meccanismi di sopravvivenza di questi miceti [20]. Inoltre, le specie del genere Candida riescono ad adattare la propria crescita alle diverse condizioni ambientali presenti nei vari distretti dell’organismo ospite. C. albicans, ad esempio, è in grado di modificare l’espressione di PHR1, un gene la cui funzione è associata alla sintesi della parete in ambiente a pH neutro (mucose del cavo orale, sangue) con quella di un gene con funzione simile, PHR2, in ambiente acido (mucosa vaginale) [21-23].
Infine, Candida può rispondere a diversi cambiamenti ambientali mediante l’attivazione di recettori cellulari di superficie che promuovono la cascata di trasduzione del segnale. Viene così modificata l’espressione genica di molti fattori di trascrizione coinvolti nell’adattamento [9,24-26].
2.1.1.3. L’invasione e la
secrezione di idrolasi.
Una volta ancorati alla superficie della mucosa, i membri del genere Candida che riescano a trovarvi condizioni idonee alle proprie necessità metaboliche, devono essere in grado di penetrare. L’invasione è favorita dalla loro capacità esclusiva di transitare dalla fase lievitiforme a quella filamentosa (morfogenesi). In generale, tale processo prevede la trasformazione delle blastospore in pseudoife, ma eccezionalmente per C. albicans e C. dubliniensis, il passaggio porta alla formazione di ife vere. Le due forme sono antigenicamente e chimicamente diverse e la forma filamentosa, dotata di capacità invasiva, è quella ritenuta responsabile dell’azione patogena [27,28].
Altri eventi che favoriscono la penetrazione sono rappresentati dall’invaginazione nella sottomucosa e dalla fagocitosi delle blastospore [29,30]. Entrambi sono promossi da adesine ed enzimi degradativi secreti.
Gli enzimi della grande famiglia delle aspartil-proteasi (SAPs, secreted aspertyl proteinases) e delle fosfolipasi (PLs, phospholipases) rappresentano i fattori di patogenicità più studiati in C. albicans [31-34]. Esistono delle differenze spaziali e temporali nell’espressione dei loro geni, ognuno dei quali sembra giocare un ruolo specifico nello stabilire un danno o nell’invadere un tessuto. Geni analoghi sono stati identificati in altre specie di Candida, come C. tropicalis, C. parapsilosis e C. dubliniensis [35-37].
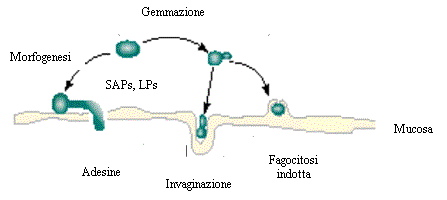
Gli eventi finora descritti, che sottintendono al processo di patogenesi, sono sommariamente riassunti nella Fig.. 2.1.1.
|
Fig.. 2.1.1. Sulla superficie della mucosa sono
rappresentati i processi di patogenesi della candidosi. Le blastospore di Candida spp. possono andare incontro a gemmazione
(al centro), oppure moltiplicarsi e penetrare nella sottomucosa promovendo la fagocitosi e
l’invaginazione (a destra). In alternativa, possono invadere i tessuti sottostanti
passando alla forma filamentosa durante la morfogenesi (a sinistra). Questi eventi sono
tutti promossi da adesine ed enzimi degradativi (SAPs e LPs). Dati modificati da Calderone
e Fonzi13.
3. La candidosi del
cavo orale.
Tra le infezioni opportunistiche che possono manifestarsi nei soggetti HIV-positivi, la candidosi del cavo orale rappresenta con certezza la patologia più frequente. Essa può preludere a forme viscerali e sistemiche che costituiscono spesso l’episodio terminale dell’evoluzione dell’AIDS.
Le localizzazioni fungine primarie o successive a disseminazione possono interessare direttamente la mucosa del cavo orale, possono coinvolgere la mucosa della lingua o le commissure delle labbra. I quadri clinici vanno dalla semplice stomatite (infiammazione della mucosa orale) alla candidosi acuta pseudomembranosa, dalla glossite eritematosa alla cheilite angolare [38,39].
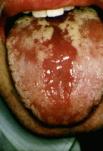
La candidosi acuta pseudomembranosa (Fig.. 3.1.a) costituisce, dopo la glossite eritematosa, la forma più comune di stomatite da Candida; essa è caratterizzata dalla comparsa in qualsiasi punto della mucosa orale, dalla bocca al faringe, di un essudato biancastro, cremoso, disposto spesso in macule a spruzzo o a chiazze e simulante un’infezione difterica. Le “pseudomembrane” se rimosse lasciano intravedere la mucosa sottostante eritematosa e dolente. Il materiale è costituito sia da componente miceliale che da cellule di desquamazione, leucociti e fibrina.
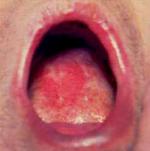
La glossite eritematosa si manifesta comunemente con lesioni piatte, sottili e rossastre, prive di essudato, di diametro variabile, sulla superficie dorsale della lingua o sul palato (Fig.. 3.1.b). Insieme alla candidosi acuta pseudomembranosa, rappresenta la prima manifestazione predittiva della transizione allo stadio di AIDS conclamato.
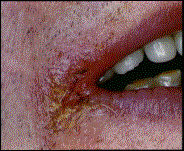
La cheilite angolare si presenta con delle lesioni a livello delle commissure labiali (Fig.. 3.1.c). Inizialmente la pelle agli angoli della bocca appare screpolata e fiacca e col progredire della malattia può andare incontro a spaccature. Le lesioni possono ulcerarsi ma l’emorragia franca è un evento raro. Le ulcerazioni non si estendono mai fino alla superficie della mucosa ma terminano sempre a livello della giunzione tra la pelle e la mucosa stessa.
a |
b |
c |
Fig.. 3.1. Candidosi del cavo orale: a) candidosi acuta pseudomembranosa; b) glossite eritematosa; c) cheilite angolare.
Circa il 80% dei pazienti HIV-positivi (prima dell’introduzione in terapia degli inibitori delle proteasi) è affetto da candidosi orale, la quale può contribuire significativamente e pesantemente nell’aggravare il quadro clinico di questi pazienti, ad esempio, nel caso in cui l’infezione si estenda dalla bocca all’esofago (esofagite da Candida). Nei casi più gravi, le complicazioni possono riguardare ascessi polmonari, endocarditi, meningiti, endoftalmiti, ascessi cerebrali, artriti e, non ultimo la candidosi disseminata, con un tasso di mortalità che può raggiungere il 70%.
3.1. L’accertamento diagnostico.
L’accertamento diagnostico di candidosi orale può essere condotto sulla base dell’osservazione delle manifestazioni cliniche e dei sintomi, soprattutto nei casi più evidenti. Molto spesso comunque, ciò può non risultare sufficiente ed occorre quindi anche un esame istologico del materiale prelevato dalla sede della lesione, osservato al microscopio grazie ad un preventivo trattamento con soluzioni di idrato sodico o potassico al 10%, che ne determinano una sufficiente trasparenza. In alternativa, l’indagine microscopica può essere effettuata dopo colorazione PAS (Periodic Acid-Schiff), ma nonostante essa possa fornire importanti indicazioni, la prova diagnostica principale resta tuttora la dimostrazione dell’agente etiologico tramite esame colturale, sia perché tale esame può risultare positivo anche in caso di negatività della ricerca microscopica, sia perché quest’ultima non fornisce, salvo eccezioni, elementi certi per l’identificazione del micete.
Per la coltura di primo isolamento da materiale patologico sono preferiti in genere, come in batteriologia, i terreni solidi. Quello di uso più comune è l’agar di Sabouraud con l’aggiunta di cloramfenicolo e cicloeximide e con un pH di 5-6. Altri terreni di coltura adoperati di frequente in laboratorio sono il Potato Dextrose agar (PDA) o lo Yeast Extract Peptone Dextrose (YEPD), ma per lo studio di peculiari caratteristiche morfo-biologiche e per una più semplice identificazione di specie si preferisce il CHROMagarÔ Candida [40,41].
3.2. La terapia.
Visto che la candidosi orale rappresenta spesso uno degli impedimenti maggiori verso il ritorno ad uno stato di salute del paziente, nel trattamento degli immunodepressi il tempestivo intervento terapeutico può risultare fondamentale [42,43].
I farmaci antimicotici comprendono sia agenti topici che sistemici. La scelta del trattamento dipende da svariati fattori, inclusi tra gli altri, il trattamento contemporaneo con altri farmaci, la presenza di alterazioni della funzionalità epatica e la tollerabilità del paziente.
Gli agenti topici includono derivati antibiotici polienici, come la nistatina e l’amfotericina B, e derivati imidazolici, quali l’econazolo, il miconazolo e il clotrimazolo, farmaci troppo tossici per essere utilizzati per via parenterale. La loro funzione è di alterare la permeabilità della parete fungina con conseguente perdita di ioni K+ e morte cellulare. Sono applicati direttamente sulle lesioni e nelle terapie a lungo termine la loro somministrazione è spesso accompagnata dall’uso di colluttori, onde evitare che gli eccipienti glucosilici in essi contenuti possano promuovere la formazione della carie.
Nei casi di infezioni profonde o di sintomi predittivi della tendenza alla disseminazione della candidosi,
come le localizzazioni oro-faringee ed esofagee, si impone la terapia profilattica
sistemica. Questa viene realizzata con la somministrazione orale di nuovi derivati
imidazolici idrosolubili: chetoconazolo, fluconazolo ed itraconazolo, scarsamente tossici
e dotati di un discreto effetto terapeutico. Nelle lesioni viscerali e di organi interni
un trattamento tradizionale è anche rappresentato dall’associazione di
5-fluorocitosina (analogo strutturale pirimidinico) con amfotericina B per via parenterale
o di amfotericina B con nistatina per via orale, sempre allo scopo di eliminare o ridurre
il focolaio micotico intestinale.
La candidosi orale tende a cronicizzare e le recidive sono frequenti, per cui può rendersi necessaria una terapia di mantenimento. In questi casi, deve essere sempre preso in considerazione anche il rischio potenziale della comparsa di fenomeni di resistenza ai farmaci [44-46].
4. L’emergenza
delle specie patogene “Candida non-albicans”.
Il cavo orale riconosce un ecologia microbica assolutamente originale e rappresenta uno degli ambienti più complessi del corpo umano, essendo capace di ospitare le popolazioni microbiche più concentrate e varie, comprese ovviamente diverse specie di Candida.
La densità della popolazione fungina cambia di frequente in base alle condizioni ambientali (dieta, trattamento farmacologico, stato di immunocompetenza, etc.) [47] e non è raro che le specie di Candida isolate da uno stesso paziente con candidosi orale siano più di una. Per cui determinare quale sia la specie che attivamente contribuisce al danno è a volte difficile.
Il principale agente etiologico della candidosi orale è C. albicans, che è comunemente riconosciuta come la specie maggiormente patogena e di più frequente isolamento fra quelle appartenenti al genere Candida [48, 49]. Nell’ultimo decennio, tuttavia, si è verificato un progressivo aumento dell’incidenza di candidosi sostenute da altre specie [50-52]. Le principali cause di tale fenomeno sono da ricercarsi nel numero sempre maggiore di soggetti immunodepressi e nei prolungati trattamenti con fluconazolo cui questi pazienti sono sottoposti. La terapia a lungo termine può infatti determinare una pressione selettiva a livello della flora fungina commensale, provocando l’emergenza di specie con ridotta sensibilità (C. tropicalis, C glabrata e C. dubliniensis) o intrinsecamente resistenti a tale molecola (C. krusei) [53-57].
La recente comparsa di C. dubliniensis come patogeno orale opportunista sembra iscriversi in questo cambiamento del quadro epidemiologico [58-61].
5. Candida dubliniensis, un nuovo patogeno del cavo
orale.
Il primo isolamento di quella che oggi è conosciuta come C. dubliniensis risale al 1957 e venne effettuato in Gran Bretagna da un prelievo autoptico di tessuto polmonare su un soggetto deceduto a seguito di una broncopolmonite [62,58]. All’epoca il ceppo isolato venne erroneamente identificato come C. stellatoidea e depositato, come ceppo di riferimento per questa specie, nella British National Collection of Pathogenic Fungi (NCPF) sotto il codice NCPF 3108. Il caso volle che Sullivan, Coleman e coll., gli irlandesi artefici della scoperta e della denominazione della nuova specie, utilizzassero questo ceppo di collezione come riferimento nella tipizzazione genetica di ceppi atipici appartenenti al genere Candida [62]. Fu così che ci si accorse della somiglianza del vecchio ceppo clinico degli anni ’50 con una serie di ceppi isolati in Australia, Irlanda e Gran Bretagna alla fine degli anni ’80 ed all’inizio dei ’90 da soggetti HIV-positivi e malati di AIDS. Solo nel 1995, tuttavia, Sullivan, Coleman e coll. proposero che questo gruppo di ceppi, fenotipicamente e genotipicamente correlati, rappresentasse una nuova specie [58]. Il nome proposto fu quello di C. dubliniensis, da Dublino, la capitale della Repubblica d’Irlanda, dove per la prima volta la nuova specie era stata identificata come tale.
Da allora molti altri isolamenti clinici di ceppi di C. dubliniensis dal cavo orale di soggetti HIV+ o malati di AIDS sono stati segnalati in Argentina [63,64], Australia [63], Belgio [65,66], Brasile [67] Canada [68], Francia [69], Finlandia [70], Germania [71], Gran Bretagna [63] Irlanda [63] Spagna [72], Stati Uniti [73,74], Svizzera [75,63]. In Italia, il primo isolamento di C. dubliniensis da campioni clinici è stato fatto a Catania dal cavo orale di un soggetto HIV-positivo [76].
La distribuzione geografica di questa specie sembra pressoché ubiquitaria, ed in un recente studio retrospettivo su ceppi clinici precedentemente identificati come C. albicans, è stata ritrovata anche in distretti corporei extra-orali, quali il tratto genitale, gastrointestinale e respiratorio, così come dai soggetti HIV-negativi [78]. In Israele C. dubliniensis è stata isolata da diversi distretti in soggetti HIV-negativi sottoposti ad antibiotico terapia [79], ed in Giappone da una ferita chirurgica [80]. Il coinvolgimento di tale specie in episodi di candidemia è stato recentemente documentato in alcuni pazienti sottoposti a chemioterapia, defedati o lungodegenti, e in corso di AIDS [81]; non è nota, tuttavia, la reale incidenza del coinvolgimento di C. dubliniensis in infezioni profonde.
I dati epidemiologici suggeriscono che, benché essa rientri fra i costituenti della normale flora microbica del cavo orale, C. dubliniensis sia in grado di causare malattia pure in assenza di concomitante infezione da parte di altre specie appartenenti al genere Candida, almeno in soggetti HIV-positivi e malati di AIDS. Il coinvolgimento di C. dubliniensis nelle patologie del cavo orale merita, poi, particolare interesse in quanto alcuni ceppi clinici sono stati trovati resistenti al fluconazolo, un antimicotico fra i più utilizzati, e ceppi sensibili al fluconazolo si sono dimostrati capaci di acquisire rapidamente resistenza a questo farmaco in vitro [82] e in vivo [83].
Dati più abbondanti sull’incidenza e la farmacoresistenza, nonché sul ruolo patogeno di questo germe, sia nei pazienti immunodepressi che nei soggetti immunocompetenti, sarebbero auspicabili per comprenderne la reale importanza. Tuttavia, perché questo sia possibile, è fondamentale che si sia in grado di identificare correttamente C. dubliniensis nei campioni clinici, distinguendola dalle altre specie del genere Candida, ed in particolare C. albicans, con la quale condivide alcuni caratteri fenotipici.
5.1. I caratteri fenotipici.
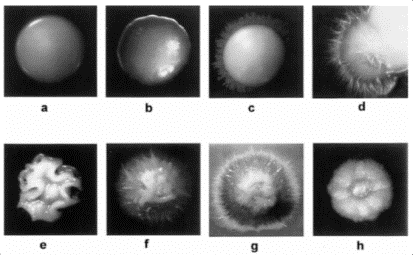
C. dubliniensis cresce bene a temperature comprese fra 30 e 37° C, sui terreni di coltura normalmente utilizzati in laboratorio per la coltivazione del genere Candida (agar di Sabouraud, PDA, etc). Le colonie appaiono cremose, di colore bianco, tonde, lisce (Fig.. 5.1.1.a), del tutto simili a quelle formate da C. albicans, con le quali condividono la caratteristica di andare incontro a cambiamenti morfologici, potendo assumere dimensioni assai ridotte, oppure aspetto rugoso o margini irregolari, con o senza la presenza di un alone opaco e sfrangiato (Fig.. 5.1.1.b,c,d,e,f,g,h) [84]. Tale fenomeno prende il nome di switching fenotipico.
|
Fig.. 5.1.1. Switching
fenotipico di C. dubliniensis e C. albicans (da Hannula e coll.84). |
Questi cambiamenti fenotipici si verificano assai di frequente in entrambe le specie, soprattutto dopo prolungati periodi di incubazione, e la somiglianza morfologica delle colonie non rende possibile differenziare le due specie nei comuni terreni di coltura. Ultimamente però l’impiego di un nuovo terreno cromogeno, il CHROMagarÔ Candida, ha permesso di ovviare a questa situazione [40]. Dopo 48 h di incubazione alla temperatura di 37° C le colonie di C. albicans su questo terreno assumono generalmente un colore verde chiaro, mentre quelle di C. dubliniensis appaiono verde scuro, risultando così facilmente distinguibili (Fig..5.1.2.a). Purtroppo, è stato osservato che col passare del tempo o a causa di ripetuti subtrapianti, le colonie di C. dubliniensis perdono la caratteristica pigmentazione [65], per cui il CHROMagarÔ Candida può essere utilizzato in maniera affidabile solo per l’identificazione delle colonie isolate cresciute dalla semina diretta del campione clinico e vanno osservate non oltre un periodo di incubazione di 48 h.
Un altro terreno utilizzabile per lo stesso scopo è il blue di metile-Sabouraud agar [65], su cui le colonie di C. albicans esposte a luce ultravioletta (lampada di Wood) producono una fluorescenza gialla, ma non quelle di C. dubliniensis. Tuttavia, anche in questo caso, i dati non sono riproducibili in seguito a vari subtrapianti o invecchiamento della colonia.
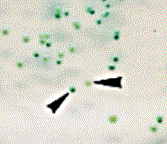
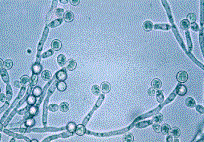
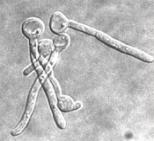
Altri importanti caratteri fenotipici di C. dubliniensis sono: la capacità di sviluppare voluminose clamidospore, sferiche, isolate o a grappoli, soprattutto quando il micete sia coltivato in terreni particolari, piuttosto poveri, come l’agar alla farina di mais (Fig.. 5.1.2.b); la capacità di formare tubuli germinativi da parte delle blastospore incubate a 37° C in siero di sangue, entro le prime due ore di incubazione (Fig.. 5.1.2.c); la formazione di pseudoife ed ife vere; la produzione di aspartil-proteasi e adesine [84]. Ma, tutti questi caratteri sono comuni ad entrambe le specie C. dubliniensis e C. albicans.
a |
b |
c |
Fig.. 5.1.2. a) Coltura primaria su CHROMagarÔ Candida
dopo 48 h a 37° C: le colonie di C. dubliniensis
appaiono di colore verde scuro (freccia sinistra), mentre quelle di C. albicans assumono un colore verde chiaro
(freccia destra); b) clamidospore apicali ed
intercalari; c) tubuli germinativi.
Tuttavia, C. dubliniensis contrariamente a C. albicans, non possiede la capacità di crescere a 45° C, di assimilare lo xilosio (XYL) e l’a-metil-D-glucoside (MDG) [73,85] e di esprimere attività b-glucosidasica [63,65,75]. E’ invece capace di ridurre il trifeniltetrazolio cloruro [86].
Le caratteristiche fenotipiche differenziali delle due specie sono brevemente riassunte nella Tab. 5.1.1.
CARATTERISTICHE
|
Candida
dubliniensis
|
Candida
albicans
|
Crescita a 45°C |
- |
+ |
Colore delle colonie su
CHROMagar Candida |
Verde scuro |
Verde chiaro |
Colore delle colonie su blue
di metile-Sabouraud agar alla lampada di Wood |
Non fluorescenti |
Giallo fluorescente |
Assimilazione di XYL ed MDG |
- |
+ |
Attività Beta-glucosidasica |
- |
+ |
Riduzione del
trifeniltetrazolio cloruro |
+ |
- |
Tab.
5.1.1. Caratteri
fenotipici differenziativi di C. dubliniensis e C. albicans.
5.2. I caratteri genotipici.
I caratteri genotipici di numerosi ceppi di C. dubliniensis sono stati studiati di recente grazie all’impiego di tecniche di tipizzazione molecolare basate sull’analisi del codice genetico.
In particolare, il confronto delle
sequenze dei geni 16S e 23S dell’RNA ribosomico (rDNA)
di ceppi di C. dubliniensis con quelle di altre
specie del genere Candida è stato utilizzato
per dimostrarne l’appartenenza ad una specie distinta [36,58]. Infatti, l’rDNA contiene regioni altamente conservate a causa dei
sui obblighi strutturali e funzionali, ma anche sequenze variabili il cui tasso di
mutazione rimane costante nel tempo (orologi molecolari) e ciò ne permette
l’utilizzo a fini tassonomici, per la definizione su base genetica di generi e
specie.
Confrontando le sequenze primarie dell’rRNA di C. dubliniensis e C. albicans, sono state riscontrate delle notevoli differenze in due regioni a forcina di un introne di gruppo I, con attività di self-splicing, dell’rRNA della grande subnunità ribosomica [68].
Il genoma di C. dubliniensis è inoltre stato studiato mediante analisi di lunghezza e polimorfismo di frammenti prodotti da enzimi di restrizione (RFLP, Restriction Fragments Length Polymorphism Analysis) su DNA microsatellite [87] ed analisi di polimorfismo del DNA amplificato casualmente (RAPD, Random Amplified Polymorphic DNA), mentre sequenze geniche specifiche di C. dubliniensis sono state amplificate grazie alla reazione a catena di polimerizzazione (PCR, Polymerase Chain Reaction) [58]. I risultati di queste analisi hanno evidenziato ancora una volta le notevoli differenze tra il genoma C. albicans e quello di C. dubliniensis. Infine, l’impiego della tecnica di elettroforesi a campi elettrici alternati (PFGE, Pulsed Field Gel Electrophoresis) [58] ha evidenziato l’esistenza di cariotipi differenti tra le due specie per la presenza di dieci o più bande di DNA, corrispondenti ad altrettanti cromosomi, in C. dubliniensis contro le sette od otto di C. albicans.
Tutti questi dati concordano nel dimostrare inequivocabilmente la natura distinta di C. dubliniensis come rappresentante di una nuova specie del genere Candida.
5.3. Le manifestazioni cliniche.
Sebbene alcuni ceppi clinici siano stati ritrovati in distretti corporei extra-orali, è soprattutto dal cavo orale di soggetti HIV-positivi ed in corso di AIDS che C. dubliniensis viene isolata più frequentemente [59].
In questi pazienti la più comune manifestazione clinica di cui C. dubliniensis è responsabile è rappresentata dalla glossite eritematosa, poiché le localizzazioni fungine primarie interessano solitamente la mucosa della lingua [62]. Tuttavia, anche localizzazioni sottogengivali sono possibili, facendo ipotizzare per questo germe un probabile ruolo nelle patologie parodontali. Infatti, analogamente a quanto evidenziato per C. albicans, anche C. dubliniensis, esprimendo fattori di virulenza in grado di determinare perdita di attacco, potrebbe essere considerata un patogeno parodontale putativo, e la sua presenza in sede sottogengivale potrebbe rappresentare un fattore di rischio per episodi di attività della malattia parodontale [88,89]. C. dubliniensis è stata inoltre recentemente implicata nella patogenesi dell’eritema gengivale lineare, un peculiare quadro di gengivite inserito fra le lesioni parodontali strettamente associate all'infezione da HIV [90].
5.4. L’azione patogena.
C. dubliniensis così come C. albicans è capace di esprimere dei fattori generalmente associati alla virulenza. Sono stati trovati ad esempio dei geni omologhi a quelli di C. albicans, sia per le fosfolipasi che per ciascuno dei 7 geni sap codificanti per le aspartil-proteasi [91,92,36,93], e solitamente i loro livelli di espressione sono risultati maggiori.
Questi enzimi rivestono un ruolo importantissimo durante la fase di attacco alla mucosa ed esperimenti in vitro hanno mostrato come l’adesione di C. dubliniensis alle cellule dell’epitelio buccale (BEC) aumenti in presenza di glucosio [36]. Questo potrebbe spiegare come mai la mucosa del cavo orale (dove la concentrazione di questo zucchero è più alta) sia maggiormente interessata alla colonizzazione di questo germe [59]. Inoltre, la capacità di C. dubliniensis di coaggregare con Fusobacteium nucleatum, un batterio del cavo orale, potrebbe favorirla nella competizione con C. albicans per la colonizzazione della mucosa orale [15].
Comunque, contrariamente a C. albicans, non sono noti in letteratura casi di un reale coinvolgimento di C. dubliniensis in infezioni profonde. Attualmente non è chiaro come possa essere limitata la sua potenziale azione invasiva ma è noto che, nonostante la forma micetale sia capace di penetrare nei tessuti più facilmente rispetto a quella lievitiforme [91], il processo di formazione dei tubuli geminativi in C. dubliniensis è più lento che in C. albicans [36]. Questo potrebbe spiegare perché C. dubliniensis sia associata solitamente ad infezioni superficiali della mucosa del cavo orale.
Infine, C. dubliniensis è capace di sviluppare più facilmente di C. albicans resistenza al fluconazolo, farmaco comunemente utilizzato per il trattamento della candidosi orale [6,56,57].
5.5. L’identificazione di C.
dubliniensis dai campioni clinici: un nuovo metodo diagnostico.
Poiché soltanto da pochi anni sono stati definiti criteri di identificazione per C. dubliniensis, non sono disponibili dati epidemiologici che permettano di verificare l'ipotesi di un ruolo parodontopatogeno per questa specie. In tale prospettiva, appare indispensabile che nei protocolli di ricerca si preveda l'utilizzo di test di laboratorio in grado di identificare correttamente C. dubliniensis rispetto alle altre specie del genere Candida, ed in particolar modo da C. albicans.
Purtroppo identificare correttamente C. dubliniensis è reso particolarmente difficile dal fatto che essa condivide con C. albicans alcuni caratteri fenotipici tra cui l’aspetto delle colonie, la morfologia delle blastospore, la formazione di tubuli germinativi in siero e la produzione di clamidospore. Per cui, la valutazione di tali caratteri, ed in particolare il Germ Tube Test (GTT), utilizzato in molti laboratori come unico criterio per l’identificazione presuntiva di C. albicans, non risulta utile nell'identificazione differenziale delle due specie. Tuttavia, la recente descrizione di metodi fenotipici, quali la valutazione del colore delle colonie cresciute sul terreno CHROMagarÔ Candida, l’incapacità di crescere a 45° C, la riduzione del trifeniltetrazolio cloruro, possono facilitare l’identificazione presuntiva di C. dubliniensis anche da campioni clinici con più specie di Candida ed anche in presenza di C. albicans. Inoltre, un’accurata identificazione può essere effettuata grazie all'utilizzo sia di sistemi commerciali che valutano il profilo di assimilazione dei carboidrati, quali i sistemi API 20C AUX ed API ID 32C [95] sia mediante l’impiego di metodi genotipici quali la PCR. Sfortunatamente però, il costo di una identificazione effettuata con sistemi commerciali risulta alquanto elevato e le apparecchiature necessarie per l’applicazione di metodi genotipici non sono alla portata di tutti i laboratori.
Il presente studio intende valutare l’utilità di due metodi fenotipici, la crescita a 45° C e la riduzione del TTC, come nuovo metodo di screening da adottare in laboratorio per confermare la presunta identificazione delle colonie di C. dubliniensis effettuata attraverso la valutazione del colore nel terreno CHROMagarÔ Candida [77].
La combinazione di questi due metodi ha permesso l'identificazione dei primi isolamenti clinici di C. dubliniensis in Italia da tamponi orali provenienti da pazienti HIV+. L’identificazione è stata confermata attraverso la valutazione del profilo di assimilazione dei carboidrati ottenuto con il sistema API 20C AUX ed i risultati sono stati confrontati tra loro per la valutazione dell’accuratezza e l’analisi costi-benefici.
L’identificazione fenotipica è stata poi confermata mediante una PCR specifica [96]. Inoltre, è stata anche accertata la sensibilità al fluconazolo degli isolati di C. dubliniensis.
Per "Materiali e Metodi", "Risultati", "Discussione" e "Bibliogafia" contattami:
 |
oppure salva in formato pdf: |
GIOVANNI M. GIAMMANCO, GIUSEPPE PIZZO, SONIA PECORELLA, SALVATORE DISTEFANO, VINCENZO PECORARO, MARIA E. MILICI. Identification of Candida dubliniensis among oral yeast isolates from an Italian population of human immunodeficiency virus-infected (HIV+) subjects. Oral Microbiology and Immunology, 2002; 17: 89-94.