
Sono piante perfettamente e completamente slegate
dall’ambiente acquatico.
Come tendenze filogenetiche, gradualmente realizzate,
troviamo:
- Predominanza dello sporofito sul gametofito, che
resta nascosto nello sporofito. Già nelle felci, lo sporofito domina sul
gametofito che è un piccolo essere di 2-3- mm., ma lo sporofito, nei primi
stadi di sviluppo, è soggetto al gametofito che lo nutre, fino a quando non
compare la prima radichetta e fogliolina. Nelle piante a fiori, è il gametofito
a essere nutrito dallo sporofito.
- Nelle felci il meristema è ad iniziale unica, ma
nelle altre piante superiori, è ad iniziale multipla, contemporaneamente
insorgono meristemi apicali e indefiniti, mentre nei muschi erano apicali ma
definiti o intercalari indefiniti; talvolta abbiamo entrambi i tipi di meristema
e la pianta, come il bambù, cresce velocemente e con forza tale da rompere i
vetri di una serra. Questo perché le piante crescono non solo in altezza, ma
anche in forza, perciò gli alberi possono rompere una roccia e il glicine
sollevare una piccola casa.
- Grazie alla comparsa di due tipi di meristema si
realizza anche un sistema vascolare non più ridotto in efficienza, come
aquixilema, ma molto più complesso.
- La pianta è così in grado di svincolarsi
dall’ambiente acqueo. Le felci si sono svincolate per quanto riguarda il soma,
ma la fecondazione avviene in acqua; nelle piante superiori l’acqua, o meglio
il liquido necessario, non è più fornito dall’ambiente ma dalla pianta
stessa.
- Si sviluppa molto anche il tessuto meccanico, in
relazione alla sollecitazioni cui le spermatophyta sono sottoposte. Questo
fenomeno si realizza in due tempi, con predominio dell’elasticità sulla
durezza o viceversa (Es: liana e sequoia).
- Graduale è l’accoppiamento fra ventilazione e
appiattimento delle foglie. Nelle Psilotopsida troviamo entità tipo foglie, ma
senza stomi, poi si giunge a foglie vere e proprie con stomi, così da aversi
massima superficie organicante e ventilazione del tessuto parenchimatico,
contemporaneamente.
- Grande sviluppo dell’apparato radicale; le piante inoltre, sanno sopperire alle deficienze ambientali, si hanno così piante grasse con radici molto lunghe dirette verso le falde freatiche e altre sottili, lunghissime, a decorso parallelo al suolo, per assorbire la rugiada della notte.
- I tessuti tegumentari si sviluppano enormemente,
sia epidermici sia peridermici, in grossi strati, tipo sughero.
- Piano piano si giunge alla completa difesa
dell’apparato riproduttore femminile, con l’Angiospermia. Tutte le piante
tendono a ciò, secondo diversi modelli, di cui alcuni hanno avuto più successo
di altri.
- La pianta è nettamente distinguibile nelle sue tre
parti: radici, fusto, foglie. Il fusto è la parte fondamentale, da cui si
dipartono verso il terreno le radici, e superiormente le foglie.
- Nella vita dello sporofito, partiamo
dall’embrione che darà la plantula, che molto presto si differenzia in
un apice vegetativo, plumula, uno radicale, radichetta e due
foglioline, cotiledoni. Tra cotiledoni e radichetta c’è una zona
intermedia chiamata ipocotile in cui si realizza il passaggio tra
struttura del fusto e della radice.
Nelle Pteridophyta abbiamo invece il piede, con cui
si assorbe il nutrimento dal gametofito.
Nel corso del tempo le parti fondamentali si
trasformano a seconda dell’ambiente. Ad esempio, nel pungitopo, le foglie sono
rami dato che portano fiori e frutti in zona centrale; le brattee della cipolla
sono foglie trasformate, che andate sottoterra, servono come riserva nutritizia;
l’Opuntia o fico d’India, le “pale” sono il fusto appiattito e le spine
sono le foglie.
Queste metamorfosi ci portano a formulare i concetti
d’analogia e omologia. I catafilli della cipolla sono omologhi alle foglie
normali, le “foglie” del pungitopo sono analoghe alle foglie normali.
La longevità delle piante è molto varia, accanto ad
esemplari plurisecolari ne abbiamo altre che vivono solo il breve periodo che
segue la pioggia nel deserto (piante effimere).
Il fusto è la parte assile fondamentale, cui si
attaccano le appendici aeree e sotterranee, le foglie e le radici; è uno sporofito.
La forma è generalmente cilindrica, ma si possono
avere forme diverse come nelle Cactacee (Opuntia, Cereus). Anche la consistenza
è molto diversificata, per cui in base ad essa le piante sono distinte in:
Erbe, con fusto non lignificato.
Suffrutici, con fusto lignificato solo alla base ed
erbaceo nella parte superiore (Timo).
Frutici o arbusti, con fusto lignificato ma con
sviluppo a cespuglio, cioè ramificato dalla base e con gemme a circa 25 cm. dal
suolo (Nocciolo)
Alberi, con fusto tutto lignificato, con un unico
asse principale che porta rami (Pino, Castagno)
La superficie può essere liscia, rugosa, bollosa,
tubercolata.
Il rivestimento può essere glabro (privo di
peli), glabriescente (qualche pelo rado), pubescente (peli corti,
poco numerosi), irsuto (peli rigidi corti), setoloso (come
l’irsuto ma con peli lunghi), lanoso o tormentoso (peli fitti
corti morbidi), appressato (peli stellati paralleli al fusto).
La direzione del fusto è molto variabile: eretto,
quando presenta una tale consistenza per cui può reggersi in modo autonomo, sdraiato
quando essendo lungo e sottile striscia al suolo (stoloni della fragola), volubile
quando riesce a sollevarsi aggrappandosi a dei sostegni (fagiolo), rampicante
quando aderisce ai sostegni mediante cirri, uncini, ventose radici avventizie
(edera), ascendente quando diventa eretto dopo un tratto strisciante.
Ci sono varie teorie e ipotesi, non suscettibili di
verifica, su come sia nato. Queste teorie le possiamo dividere in due grandi
categorie.
I ) Il fusto è la parte fondamentale che si ramifica
nel corso del tempo.
II) La foglia è la parte fondamentale.
Secondo le prime teorie, sorge ora il problema di
come sia nata la foglia. Le possibilità sono due:
I ) appendici cilindriche, e cioè i rami si
sarebbero saldati, dando un qualcosa di laminare. Si può ammettere ciò per la
Rhynia; è un processo chiamato di cladodificazione.
II) fra un ramo e l’altro si forma una lamina, tipo
zampa palmata dell’anatra in un processo chiamato laminazione. Secondo
altre teorie, il fusto sarebbe nato dalla saldatura dei piccioli delle foglie.
Quale sia la vera morfogenesi, non si sa; mentre della prima teoria abbiamo un esemplare nella Rhynia che ha fusto e rami ma neppure una foglia, per cui queste sono certamente comparse dopo, della seconda teoria non si ha prova alcuna.
Stabilito che il fusto è la parte fondamentale, consideriamo la teoria del teloma, secondo cui il fusto è la parte più primitiva della pianta, che si ramifica in alto dicotomicamente, una volta sola; qualora il ramo nato da una dicotomia, si dicotomizzi a sua volta, si parla di sinteloma. È una teoria che trova conferma nella Rhynia, fossile.
Da un teloma si potrà avere solo una foglia, per saldatura dei due rametti; da un sinteloma, le ultime dicotomie si salderebbero a dare più foglie.
La Pteridophyta, Psilotum triquetum, è un sinteloma e potrebbe essere derivato dalla Rhynia, ma mentre di quest’ultima abbiamo molti fossili e nessun esemplare vivente, per lo Psilotum è esattamente il contrario.
Il fusto prende origine da una segmentazione di meristemi e precisamente da quella parte dell’embrione da noi chiamato apice vegetativo. Il meristema può essere ad iniziale unica o ad iniziale multipla e in quest’ultimo caso, le cellule apicali dell’apice vegetativo daranno origine a tutte le possibili parti del fusto. Piano piano il fusto si ramifica e all’apice dei rami troviamo porzioni di apice vegetativo, cioè meristemi primari.
Ma c’è un nuovo problema; quali cellule dell’apice daranno origine ai rami e quali ad altre parti del fusto?
Logicamente è un problema di difficile soluzione e il difetto delle varie teorie è sempre quello di voler estendere a tutti i vegetali risultati che hanno avuto conferma sperimentale solo per alcuni di essi.
Tra queste teorie, ricordiamo quella degli istogeni e quella della tunica-corpus.
Istogeni: nel meristema ad iniziale multipla abbiamo tre gruppi di cellule che danno origine alle diverse parti del fusto; dall’esterno all’interno, troviamo, un dermatogeno che origina l’epidermide, un periblema che dà la parte corticale, un pleroma che dà i tessuti conduttori. Questa teoria non regge per varie ragioni e opportune ricerche hanno dimostrato che le cellule dei meristemi, dividendosi non in senso periclinale ma anticlinale, non darebbero possibilità di ramificazioni laterali.
Tunica corpus: qui ci sono due gruppi di cellule, uno dante la tunica e l’altro il corpus; la tunica sarebbe tutta la parte periferica della pianta e il corpus solo la parte centrale, cioè il midollo. Così nell’apice vegetativo avremmo una parte centrale dante il corpus e una periferica dante la tunica. Uno degli assurdi, sta che mentre la tunica darebbe tutti i possibili tipi di tessuto, il corpus darebbe solo un midollo di natura parenchimatica.
La teoria più accreditata secondo studi più recenti è la seguente: nell’apice vegetativo ci sarebbero tre tipi di meristema, protoderma, procambio dante i tessuti vascolari, meristema fondamentale dante i tessuti parenchimatici. Essi non sarebbero localizzati topograficamente in regioni distinte, ma le cellule degli uni e degli altri sarebbero frammiste, e cioè una cellula darà l’epidermide e quella immediatamente vicina darà del tessuto vascolare.
Quando l’apice vegetativo si divide in tante porzioni, avremo tanti apici vegetativi, il che comporta distinguere l’apice vegetativo del fusto vero e proprio da quello di ramificazioni laterali. Avremo poi anche periodi di arresto dell’attività moltiplicativa.
Indichiamo come gemma, qualunque parte della pianta provvista di meristema apicale e con apice vegetativo. Queste gemme sono qualcosa che già in potenza sono un ramo intero, dato che in esse non troveremo solo l’apice vegetativo che darà il ramo centrale, ma anche tutti quegli apici che daranno rami laterali e foglie, per cui una gemma è definibile come un insieme di meristemi già differenziati che daranno le diverse parti.
Sezionando una gemma, troveremo all’apice, il vero apice vegetativo del ramo centrale e poi, lateralmente, una serie di bozze, alcune più piccole, altre più grandi, le più piccole rappresentano i primordi dei rami, le altre sono le foglie. Le gemme possono essere ascellari o apicali. Mentre il meristema nelle gemme dei rami è indefinito, quello delle foglie è definito e darà origine all’apice precursore.
Tutto questo complesso è protetto dalle perule, squamette di colore marrone, talvolta glutinose, ben visibili nell’ippocastano.
Talvolta l’abbozzo del ramo ha un meristema apicale ad accrescimento definito, questo avviene quando il ramo darà origine all’apparato sporigeno, cioè al fiore; troveremo questa condizione in quelle che noi chiamiamo gemme fiorifere, in cui i primordi dei rami sono bloccati.
Le gemme sono ridotte nelle regioni equatoriali, in cui le piante si trovano in condizioni ottimali tutto l’anno, mentre sono grandi ed efficienti in quei paesi in cui l’inverno ha una sua precisa fisionomia.
Il fusto si presenta con una
parte assile detta in genere tronco o asse o fusticino, in genere caule.
Notiamo nel caule i punti di partenza dei rami e in queste zone degli ingrossamenti detti nodi, mentre la zona fra nodo e il successivo, è detta internodo ed in genere qui, il fusto ha diametro minore che nel nodo, dov’è anzi ingrossato. Dal nodo parte un ramo di primo ordine e dai nodi di questo, rami di secondo ordine, e così via. All’apice del ramo e del fusto troviamo un centro meristematico o apice vegetativo di diversa importanza.
Molti fusti presentano anche accrescimento intercalare, perché l’apice lascia via via dietro di sé delle cellule meristematiche che proliferano per un tempo definito e costituiscono un meristema intercalare, in genere sopra al nodo. Nel bambù, l’accrescimento è notevole e in pochi giorni, può crescere di 4-5 cm. grazie ai meristemi apicali e intercalari che possiede.
La ramificazione avviene in modo diverso da specie a specie. Può essere dicotomica, quando l’asse si divide in due parti o perfettamente simmetriche (isodicotomia) o no (anisodicotomia). In questo caso il ramo che si accresce può essere il secondario, per cui l’apice del fusto è più in basso di quello del ramo (ramificazione simpodiale) o viceversa (ramificazione monopodiale). Nella ramificazione monopodiale, l’asse continua ad accrescersi emettendo rami laterali, è caratteristica delle Conifere (abete, cipresso, ecc.); quando invece, l’attività della gemma terminale cessa, mentre continua lo sviluppo dei rami laterali, si parlerà di ramificazione simpodiale ed è caratteristica delle Dicotiledoni (melo, nespolo, noce, platano, ecc.).
La più antica dicotomia è l’isodicotomia, ed è in relazione con un meristema ad iniziale multipla proliferante in modo equivalente nelle due metà. Se il meristema è ad iniziale unica, è molto difficile avere un sistema del genere, perché le tre facce si accrescono con diversa velocità ed avremo quindi, sempre un’anisodicotomia.
Nella monopodiale, l’apice vegetativo prolifera in tre direzioni, due laterali e una centrale; se prevale la proliferazione centrale avremo una ramificazione monopodiale, se prevale la proliferazione laterale, così da superare la centrale, avremo la ramificazione simpodiale.
In relazione a questo fatto, nella simpodiale, l’attività della parte centrale è minore nella stagione cattiva, mentre nella monopodiale prolifera costantemente tutto l’anno, quindi la simpodiale è propria delle piante che in inverno perdono le foglie, la monopodiale delle piante sempre verdi.
Per quanto riguarda l’anatomia del fusto, lo studio delle sezioni ci ha portato a fare delle distinzioni. Quindi, nel fusto, è necessario distinguere, una struttura primaria, del primo anno di vita, ed una secondaria che insorge a due anni e si mantiene per tutta la vita. La struttura primaria, si mantiene tutta la vita anche nelle monocotiledoni, mentre tutte le altre piante passano alla struttura secondaria, come anche certe felci e certe conifere, oltre che tutte le dicotiledoni.
La struttura primaria è solo la struttura del periodo vegetativo, perché anche in quelle piante che non la cambiano, con il sopraggiungere della brutta stagione, si ha il comparire di una struttura di riposo vera e propria, o almeno, di una qualche variante alla precedente anatomia.
Esaminando dall’esterno la sezione, cilindrica, di un fusto in struttura primaria, troviamo, l’epidermide tappezzante e chiudente il cilindro completamente, provvisto di stomi. Quest’epidermide è originata dal meristema detto protoderma, che proliferando, forma tante cellule e tappezza tutta la parte periferica del cilindro. Generalmente è monostratificata, (può essere anche bistratificata) fatta da un solo strato di cellule allungate tangenzialmente, con la parete esterna molto indurita, in quanto chitinizzata.
Troviamo poi uno strato di corteccia o strato corticale, prevalentemente fatto di tessuti parenchimatici accoppiati a tessuti meccanici, che deriva dal meristema fondamentale che dà origine alle cellule parenchimatiche. Sotto di questo troviamo un anello di collenchima dai caratteristici ispessimenti angolari e anche isole di sclerenchima, ma mai uno strato continuo c’è anche del parenchima clorofilliano fatto da cellule ricche di cloroplasti, in cui si compie l’organicazione del carbonio; in genere questo strato fototrofico è più esterno e il meccanico più interno, ma senza una netta distinzione far i due. Più internamente ancora troviamo parenchima amilifero di riserva. Sotto di questo, uno strato detto endoderma, che non si può più chiamare corteccia e ha una funzione analoga a quella dell’epidermide e separa la corteccia dalla parte centrale. Si può trovare conferma di ciò, nelle piante più antiche, dove l’endoderma va a disporsi attorno ad ogni singolo fascio vascolare, spingendosi sino al midollo. Nella radice è molto più evidente, per avere le cellule ispessimenti ad U, mentre nel fusto, si evidenzia molto bene quando si presenta come uno strato unico di cellule allungate tangenzialmente. Queste cellule presentano, nelle parti radiali di divisione degli ispessimenti in zona centrale, detti strie di Caspary. Talvolta, queste cellule dell’endoderma si stipano di granuli d’amido in modo abnorme, si parla allora di guaina amilifera all’interno dell’endoderma, troviamo il cilindro centrale o stele, costituito fondamentalmente dalla parte vascolare. Quindi abbiamo, una parte periferica detta periciclo, fatta da tessuto parenchimatico di riserva, più internamente i fasci vascolari e fra un fascio e l’altro, i raggi midollari parenchimatici. Più internamente, al centro della sezione, c’è il midollo, tessuto parenchimatico variamente differenziato e talvolta lignificato.
Il periciclo è sempre fatto da parenchima di riserva, mai clorofilliano, perché lontano dalla luce e la riserva è in genere amilifera. Può essere fatto da 1 a 4-5-6 strati di cellule, comunque sempre un numero basso. La zona vascolare, in struttura primaria, è fatta da fasci cribro-legnosi di vario tipo:
1 – fascio collaterale chiuso con floema esternamente e internamente lo xilema a diretto contatto,
sovrapposti.
2 – fascio collaterale aperto con floema e xilema e interposto fra essi uno strato di cambio,
meristema primario.
3 – fascio bicollaterale con xilema intervallato fra due zone di floema. Proprio di piante con fusto
cavo, tipo le zucche.
4 – fasci concentrici con floema esternamente e xilema internamente, detti perifloematici.
5 – fascio concentrico perixilematico con xilema esternamente e floema internamente.
6 – fasci con floema e xilema intervallati e separati, ma incontrantisi al centro, detti fasci alterni a
struttura alterna, caratteristica della radice.
Considerati i fasci, è ora necessario vedere le mutue posizioni di proto- e meta-xilema, mentre non si considera il deuteroxilema, in quanto comparente nella struttura secondaria.
Il protoxilema si forma nei primi momenti di formazione del fascio, mentre il metaxilema si forma ad opera del cambio. Quando il protoxilema è all’interno del fascio, si dice endarco, se esterno, si dice esarco, se interno mesarco.
Questi fatti sono importanti perché nel fusto, in genere, il protoxilema è endarco, mentre nella radice è mesarco, ci sarà quindi una zona di passaggio da una all’altra struttura. Questo avviene nell’ipocotile, cioè in quella parte dell’embrione che sta sotto l’apice vegetativo e prima della parte radicale; nell’adulto, detta zona è individuabile nella parte del fusto immediatamente sotto l’inserzione dei cotiledoni, fino a livello terra. È sempre in questa zona che avviene il passaggio dalla struttura alterna della radice a quella del fusto.
La stele non è altro che il fusto. È difficile dimostrare la derivazione da un tipo all’altro di stele, poiché molti esemplari sono fossili e mancano effettivi termini di passaggio.
Consideriamo il disegno della pagina 27. Attorno ad ogni disegno, dobbiamo immaginare un altro cerchio nero che rappresenterebbe il margine del fusto e sotto di questo una zona bianca che rappresenta tutto ciò che è compreso fra epidermide ed endoderma. Il cerchio nero, qui raffigurato, indica appunto l’endoderma; scompare nelle steli in cui l’endoderma si spezzetta a circondare singolarmente i singoli fasci.
Il floema è rappresentato tratteggiato, lo xilema scuro. I disegni numerati 12 13 14 15 16 rappresentano tutte le steli a protoxilema esarco; le altre hanno protoxilema endarco o mesarco, le frecce indicano la derivazione.
Usiamo queste abbreviazioni: F = floema ; X = xilema ; PX = protoxilema
1) Protostele : F. esterno, X. centrale, PX. Endarco; concentrici; un solo endoderma. Da questo
tipo di stele, prendono origine due linee evolutive, una con PX endarco o
mesarco, l’altra con PX esarco; da quest’ultima per midollazione origina la
2) Sifonostele in cui al centro compare una massa midollare. Da questa deriva la
3) Sifonoeustele in cui la massa di X si è divisa in tante porzioni.
4) Eustele in cui si è frammentato pure il F. e abbiamo tanti fasci ben separati (dicotiledoni
e conifere) con endoderma unico.
8) Polistele derivante da sifonoeustele. L’endoderma si è spezzettato attorno ai diversi fasci
con PX. Endarco (felci).
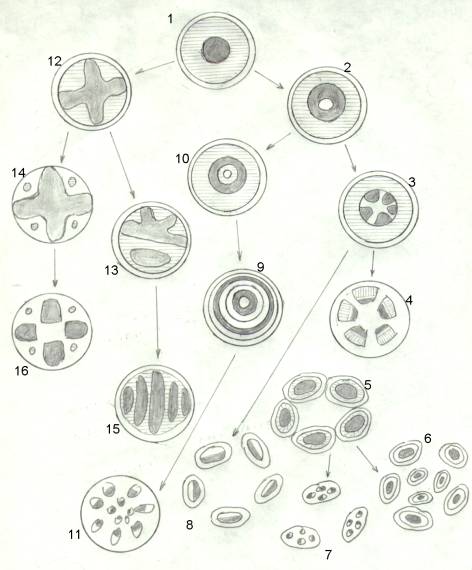
1 - protostele 2 - sifonostele 3 - sifonoeustele 4 - eustele
5 - dictiostele 6 - polidictiostele 7 - polieustele 8 - polistele
9 - policiclostele l0 - sifonostele anfifloica 11- atactostele 12 - protostele stellata
13 - plectostele 14 - actinostele piena 15 - aciclostele 16 - actinostele midollata
10) Sifonostele anfifloica : deriva da sifonostele. Il PX è rimasto chiuso fra due masse
moltiplicatisi in senso inverso, cosicché abbiamo, in successione,
F-X-PX-X-F in cerchi concentrici, con endoderma unico.
11) Dictiostele : deriva dalla precedente. PX mesarco e molti endoderma, ognuno circondante il
fascio, ogni fascio, costituisce un’entità detta meristele.
6) Polidictiostele : in cui le diverse meristele si sono moltiplicate su cerchi concentrici.
7) Polieustele : deriva dalla precedente; più fasci hanno lo stesso endoderma.
9) Policiclostele : deriva da sifonostele anfifloica. Fra le porzioni di F e X si forma un cerchio di
parenchima midollare. Il PX ritorna endarco e compare in ciascun cerchio.
11) Atactostele : sorgono raggi midollari fra fascio e fascio, per cui i fasci si spezzettano in tanti
più piccoli, concentrici, isolati, con endoderma unico (monocotiledoni)
Dalla protostele possono prendere origine pure le steli con PX esarco
12) Protostele stellata : Lo X è a stella, con agli apici il PX.
13) Plectostele : perdita completa della simmetria (Licopodiaciee)
15) Aciclostele : l’endoderma è esterno; lo X si è organizzato in tante lenti, ciascuna con il suo PX
esarco (Selaginelle).
14) Actinostele piena : struttura alterna con fasci di solo F e altri di solo X; deriva da protostele
stellata.
16) Actinostele midollata : deriva dalla precedente, per frammentazione dello X e successiva
midollazione centrale.
Lungo il fusto s’inseriscono le foglie, anch’esse provviste di tessuto vascolare. Anatomicamente, cosa avviene?
Un pezzetto di fusto, estroflettendosi, va a costituire la traccia fogliare. In corrispondenza di tale distacco, abbiamo un’interruzione nella parte vascolare, dato che questa è andata a costituire la foglia ed è detta lacuna fogliare, di natura parenchimatica, senza più tessuto vascolare.
Si può notare dal disegno, come lo xilema sia sempre interno e il floema esterno.

Nella sezione B, è rappresentata la situazione quando si è realizzato il distacco dal fusto di foglie e rami. In genere gli abbozzi dei rami sono due, mentre uno solo è quello della foglia, che qui presenta anche i primi abbozzi della nervatura, fatti per metà da floema e per metà da xilema.
Mentre la traccia fogliare, ha sempre andamento periclinale, cioè segue la generatrice del cilindro, le tracce dei rami hanno andamento anticlinale, cioè come un raggio che passa per il centro. Nella foglia c’è poi l’impossibilità di formare un cerchio, mentre nel ramo, per proliferazione laterale delle tracce, si realizza una struttura circolare chiusa (passaggio da A a B).
Le tracce dei rami sono sempre due, in quanto si formano ai lati della foglia. Talvolta le cose sono ancora più complesse perché abbiamo più tracce fogliari; nel bambù, le tracce fogliari sono molteplici, per cui avremo un sistema multilacunoso. Tutta questa complessa struttura, si realizza nel punto detto nodo. Quanto descritto è lo schema generale e normale, col ramo in posizione superiore e la foglia sotto, possiamo però, avere posizioni invertite e anche le due entità su differenti ortostiche, intervallate col ramo a livello più basso (Equisetum).
Nelle monocotiledoni, la struttura resta sempre primaria, ma si ha la comparsa di nuovi fasci per la presenza di un meristema a gradino. Il fascio è chiuso, cioè senza una zona intermedia di cambio; il procambio si è esaurito nel dare il fascio e non ha potuto avere la forza sufficiente a dare cellule meristematiche interposte, cioè quelle cellule che sono le dirette responsabili della struttura secondaria degli altri gruppi di vegetali.
Logicamente, la struttura secondaria non insorge di
colpo, ma si ha un passaggio graduale.
Nelle dicotiledoni e nelle conifere, si ha in genere
una eustele di fasci di tipo aperto. Al momento del passaggio alla struttura
secondaria, insorgono due anelli concentrici di meristema, il fellogeno e il
cambio. Il cambio prolifera, da entrambe le parti, in senso periclinale, cioè
parallelamente alla superficie. In questo modo, nella zona del raggio midollare,
compaiono cellule meristematiche e i due bracci di cambio provenienti da fasci
distinti, finiscono col congiungersi, proprio in detta zona, al centro; è
comparsa così, una zona meristematica, che deriva da cellule parenchimatiche
del raggio midollare. Viene a realizzarsi, in questo modo, un intero cerchio di
cambio secondario e dentro i fasci troviamo un cambio intrafasciale derivante da
meristema primario, mentre il cambio interfasciale deriva da meristema
secondario; siamo, ora in presenza di una struttura intermedia tra quella
primaria e quella secondaria.
Le cellule che hanno riacquistato proprietà
meristematiche, sono quelle del raggio midollare che si trovano a contatto col
floema; ciò è importante perché si ritiene che sia proprio il floema a dare
la spinta in questo senso, grazie alle sostanze nutritizie che porta, perciò si
può dire che il floema è l’organizzatore di detto cambio. Questo cambio, sia
l’intrafasciale sia l’interfasciale, comincia immediatamente a proliferare,
appena formato, dando floema verso l’esterno e xilema verso l’interno. La
modalità di moltiplicazione è la seguente: una cellula si divide
tangenzialmente in due, ma di queste una sola differenzia il floema e l’altra
resta meristematica. Questa poi, si divide ancora e, una dà lo xilema, e
l’altra resta meristematica e così via, con alterna formazione di
deuterofloema e deuteroxilema. Ciò comporta un gioco d’anelli concentrici
anche all’interno dei fasci, allontanando sempre più le parti primitive di
floema e xilema. Ogni anno compare un nuovo anello, da cui la possibilità,
contandoli, di determinare l’età dell’albero.
Sotto l’epidermide, si origina il fellogeno che, a
differenza del cambio che dura tutta la vita, si squama gradualmente e sempre di
nuovo se ne forma; è perciò un meristema a gradino (siamo in struttura
secondaria). Variabile è la sua localizzazione, più o meno profonda e in
definitiva, è come un anello concentrico esterno a quello del floema; ogni anno
si forma un anello di floema, xilema e, più esternamente, fellogeno che
progressivamente desquama in placche di ritidoma.
Considerando la sezione di un fusto e osservando dall’esterno verso l’interno, troviamo sughero o fellema, fellogeno e felloderma, poi la corteccia, la zona vascolare col floema del fascio primitivo, floema dell’anello, cambio, xilema dell’anello, xilema primitivo.
I cerchi concentrici, verso l’esterno si rompono, in quanto il fusto si accresce. Compaiono, così, dei cunei dovuti a cellule parenchimatiche e costituenti i raggi midollari. Allorché la corteccia secondaria si distacca, ecco insorgere nuovo fellogeno e nuovo sughero, immediatamente sotto la vecchia corteccia, a protezione dei tessuti più interni, isolandoli così dall’ambiente esterno. La corteccia si distacca in placche di ritidoma di varia forma, a fogli, a placche, a zone, a coste ecc. spesso si desquama anche il floema, qualora il fellogeno si formi troppo internamente.
Per quanto riguarda la durata nel tempo delle porzioni costituenti la struttura secondaria, si osserva che: come sopra detto, la corteccia secondaria si distacca come ritidoma, lo xilema resta sottoforma di anelli concentrici che aumentano di numero ogni anno. Il cambio forma anelli in relazione alle stagioni; nelle zone equatoriali, gli anelli non sono ben distinti, perché gli elementi xilematici sono tutti uguali, data l’uniformità del clima tutto l’anno; nei climi temperati, dove esiste una differenza stagionale evidente, con un periodo invernale di riposo della pianta e quindi arresto della sua attività e caduta delle foglie, alternato ad un periodo estivo di forte attività, il cambio cessa di lavorare in inverno e riprende solo il suo funzionamento in primavera. In questo periodo forma degli elementi xilematici molto più grossi, in relazione all’abbondanza di acqua; con l’avanzare della stagione e la mancanza progressiva di acqua, gli elementi xilematici diminuiscono le dimensioni, fino alla cessazione del loro funzionamento. Abbiamo così, un legno di primavera e uno estivo; nell’anello le diverse dimensioni degli elementi sono ben visibili per cui un albero ci fornisce anche utili indicazioni climatiche. In un albero di quattro anni è possibile vedere le parti residue, in genere cuneiformi, del floema e xilema primitivo. Nel fusto, questi residui, sono sullo stesso asse mentre nella radice sono su assi distinti, cioè in una struttura alternata. Ciò permette di riconoscere un giovane fusto da una radice.
Talvolta questo legno secondario è nettamente differenziato in due zone diversamente colorate, ciò è in relazione al fatto che il legno cessa ad un certo punto di funzionare come tale e diventa elemento di sostegno assumendo un colore scuro e dicesi duramen, mentre la parte più esterna, conducente e più chiara, è detta alburno.
Nelle monocotiledoni plurienni, si forma una zona di accrescimento costituita da cellule parenchimatiche che non differenziano mai in meristema secondario vero e proprio, ma semplicemente differenziano nuovi fasci e non si raggiunge mai una struttura secondaria. È un meristema a gradino, che si forma sempre più esternamente a quello che ha cessato di funzionare; si differenzia così nettamente dal fellogeno che, quando si riforma, si localizza sempre più addentro, lasciando fuori la parte morta, ossia il ritidoma.
Il fusto si trasforma, adattandosi all’ambiente. Le principali modificazioni sono le seguenti:
Rizoma: Es. Iris (Iris germanica). È un fusto ipogeo, ossia sotterraneo, che è diventato dorso-
ventrale e cioè mette radici solo nella parte volta verso il basso. La parte vascolare è
ancora discretamente sviluppata mentre molto di più lo è la corteccia. È protetto dalla
terra che lo ricopre ed è praticamente un magazzino di riserva.
Tubero: Es. Patata (Solanum tuberosum). Il fusto non solo è sotterraneo ma anche profondamente
infossato; ha simmetria raggiata ed è un abbondante magazzino di sostanze di riserva; il
tessuto vascolare è ridotto al minimo e abbondante è il parenchima amilifero.
Bulbo: fusto ipogeo formato da un “cuore” detto girello con gemma terminale e con catafilli
(foglie trasformate) più o meno carnosi. Si distinguono in:
Bulbo tunicato: Es. Cipolla (Allium coepa). Il fusto vero è il girello. È un fusto che si è
estremamente appiattito ed ha un estremo accorciamento degli internodi, per
cui le foglie (catafilli) sono strettamente unite. Se i catafilli sono embricati, cioè
sovrapposti in modo alternato, il bulbo dicesi squamoso; se i catafilli coprono in
modo completo quelli interni, dicesi tunicato. L’aglio (Allium sativum) ha un
bulbo composto.
Bulbo solido; Es. Colchico (Colchicum autumnale). I rami e la base del fusto si sono trasformati
in sostanze di riserva.
Fusti formicai: in certe piante, la base del fusto si è ingrossata molto ed è fatta da tubercoli con
gallerie in cui vivono le formiche in simbiosi (mirmecofilia). Le formiche
accumulano qui le proprie provviste e le piante ne utilizzano una parte. Le pareti
di questi tubercoli sono lisce rivestite d’epidermide.
Stoloni: Es. Fragola (Fragaria vesca). I rami corrono paralleli al terreno per un certo tratto,
emettono radici e nuove pianticelle, che successivamente rompono il collegamento
con la pianta madre; propagazione.
Fusti succulenti: Es. Fico d’India (Opuntia ficus indica). Tutte le piante grasse hanno questo tipo
di fusto. C’è abbondanza di tessuto acquoso e una fitta rete di tessuto vascolare.
Cladodi: Es. Pungitopo (Ruscus aculeatus). Quelle che comunemente chiamiamo foglie, sono in
realtà i rami appiattiti (cladodi), dato che su di essi, nella parte inferiore, sboccia il fiore.
Sono in grado di compiere la funzione clorofilliana.
Il nome del Pungitopo deriva dal fatto che con esso si facevano grandi trecce, i gaschi,
che posti attorno alle gomene, impedivano ai topi di salire sulle navi.
Spine: Es. Biancospino (Crataegus oxyacantha) , Prunus. Le spine sono provviste di vasi e
staccandole si producono lacerazioni nei tessuti sottostanti. Le spine delle rose, sono foglie
trasformate e facilmente si staccano.
Turione: Es. La parte edule dell’asparago. E’ un germoglio con foglie appena accennate e scarsa
clorofilla.
Viticci: Es. Vite (Vitis vinifera). Sono apparati di presa, da non confondere coi cirri dei piselli, che
derivano dalle foglie.
Viticci adesivi: Es. Vite americana, (Parthenocissus). Sono apparati di presa che si attaccano
per mezzo di piccole ventose e quindi non circondano il supporto.
Secondo la posizione delle gemme e del portamento, si stabiliscono fra le piante delle forme biologiche. Nelle zone tropicali, le piante sono generalmente alte e di grandi dimensioni, nelle zone di alta montagna, le piante sono di dimensioni minori o addirittura si ha la scomparsa di esse. Nella stessa zona è inoltre possibile vedere comportamenti diversi. Esistono piante con gemme non protette, altre protette. Quindi distinguiamo diverse forme:
Camefite : con gemme sotto i 25 cm. dal suolo.
Suffrutici vari (Thymus, Alyssum).
Emicriptofite :
con gemme sul piano del suolo. Sono cespitose come l’erba, reptanti come la
fragola, rosolate con rosette di foglie, come la Bellis. Sono gemme
protette dai soli
detriti fogliari dell’anno prima.
Neofite: con gemme sotto terra, perché sono portate da
rizomi e bulbi, sopportano i geli invernali.
Terofite: sono piante che passano la stagione avversa come
semi, o già come pianticelle (grano) e
germinano solo con la stagione favorevole. Alcune sono effimere, vivono
per poco
tempo ma sono in grado di fiorire e fruttificare nel breve periodo di
pioggia, come
avviene nei paesi desertici.
Idrofite: rizomatose, vivono ai margini degli stagni (Typha).
radicanti, hanno radici interrate e foglie che galleggiano come le
ninfee.
natanti, come l’Eichornia crassipes, infestante o la Pistis stratiotes,
simile ad un cespo
d’insalata, dalle foglie spesse, propria dei tropici.
Lianose: poche le liane presenti da noi (Vitalba).
Epifite erette: caratteristico è il vischio.
Nei climi tropicali predominano le Fanerofite, grandi; ove abbiamo un’alternanza tra stagione secca e piovosa, abbondano le Nanofanerofita, le emicriptofita e le Microfanerofita; nei nostri climi, ove caratteristica è la macchia mediterranea, prevalgono gli arbusti; verso nord, cioè verso le zone alpine, le Camefite. Nei luoghi palustri, c’è abbondanza di Idrofite, nei paesi tropicali, molte sono le Epifite, mentre da noi ci sono solo muschi e licheni.
La foglia è un’appendice del fusto, ovvero la parte laminare destinata a tre funzioni: traspirazione (eliminazione d’acqua allo stato di vapore), respirazione (assunzione di ossigeno ed eliminazione di anidride carbonica), fotosintesi clorofilliana (complesso fisico-chimico che utilizza l’energia della luce per organicare il carbonio). Di grandezza variabile, non solo nelle diverse specie ma anche sullo stesso individuo; variabile nelle diverse specie ma costante nelle singole specie è invece la forma. È percorsa da nervature.
Cinque sono i tipi di foglie che si succedono lungo
il fusto dal basso verso l’alto: i cotiledoni o embriofilli, con
funzione nutritiva e protettiva dell’embrione; i catafilli con funzione
di protezione delle gemme e di riserva; i nomofilli, rappresentati dalle
tipiche foglie, con funzione di traspirazione e sintesi clorofilliana; gli ipsofilli
per la protezione del fiore; gli antofilli con funzione riproduttiva. Le
prime foglie dopo i cotiledoni, vengono dette profilli.
La foglia è essenzialmente formata da: lamina o lembo, picciolo, guaina, stipole. Queste tre ultime possono mancare.
Secondo la teoria del teloma e del sinteloma, la
foglia nasce per cladodificazione, cioè saldatura di più rami o per
laminazione, cioè per l’insorgenza di una membrana fra i rametti. Cosa, però
fondamentale, è la dicotomizzazione, in quanto è il primo fatto che si è
verificato e senza il quale, le foglie non si sarebbero mai formate, perché
solo con la dicotomia sono comparsi quei rami fra i quali può essere avvenuta
una cladodificazione o una laminazione.
Le foglie più primitive sono quelle che derivano da
cladodificazione, dato che con questo sistema le unità fogliari non saranno mai
molto espanse essendo i rami limitati di numero; le foglie più moderne sono
quelle derivanti da laminazione.
Per quanto riguarda la morfogenesi, è necessario
considerare che il fusto, ha all’apice una zona meristematica in cui una parte
centrale provvede all’accrescimento del fusto e lateralmente porta le bozze
fogliari. Mentre però il meristema del fusto è indefinito, quello delle foglie
è definito, bloccato. Esiste però una sorta di compenso: mentre il meristema
del fusto si accresce lentamente per tutta la vita, quello della foglia è un
accrescimento rapido, ma ad un certo punto muore.
Alla nascita,
la foglia ha un piccolo meristema apicale che dà un limitato numero di cellule
e poi cessa di funzionare. Questo meristema è sostituito da un meristema basale
intercalare che velocemente dà cellule in senso basipeto, formando, nella
successione temporale e spaziale, lamina, picciolo, guaina, stipole. A questo
punto, la foglia si è completata e cessa di crescere.
Il meristema basale intercalare produce sempre nuove
cellule verso la base, le cellule più vecchie della foglia sono quelle apicali,
a contatto con l’apice precursore, mentre le più giovani sono a contatto con
le stipole. Questo accrescimento è anche allometrico, cioè diverso nelle tre
direzioni dello spazio, minimo per lo spessore, massimo in lunghezza, minore in
larghezza. L’accrescimento è anche pleuroplastico, cioè un accrescimento non
solo al centro ma anche laterale.
Se prevale l’accrescimento pleuroplastico, abbiamo
una foglia reniforme se prevale l’acrocarpico abbiamo una foglia allungata. È
proprio l’accrescimento particolare del meristema a determinare quella che sarà
la forma della foglia. Esistono esempi rarissimi di foglie con meristema basale
indefinito. Tale la Welvitschia, pianta con un tronco di 2-3 m. di diametro, ma
con due foglie laminari, opposte che si accrescono indefinitamente.
Un altro tipo di foglie interessante è quello delle
felci. Più propriamente, dovremmo chiamarle fronde, tenendo conto che
sono in realtà sporofilli, cioè “foglia” portanti su di sé gli apparati
sporigeni. Hanno in genere accrescimento definito, ma in Licodium, ad esempio,
il meristema è apicale indefinito, fatto questo importante, a favore della tesi
secondo cui le “foglie” delle felci sarebbero rami. Ipotesi questa,
confermata anche dall’anatomia, infatti, qui troviamo, endoderma e meristeli.;
crescono per 4-5 anni, partendo dalla forma a “pastorale” e svolgendosi
piano piano.
La foglia prima si forma, poi si apre. Il primo
processo è detto di prefogliazione, cioè lo stadio precedente alla
stiratura. La prefogliazione può essere: circinnata (a ricciolo), placata
(a fisarmonica), diritta ecc. Già nella gemma possono avere questo
aspetto.
La morfologia definisce tutte le caratteristiche
della foglia e comprende: l’inserzione o fillotassi, la direzione,
l’attaccatura al fusto, la nervatura, la forma della lamina, il margine della
lamina, la base della lamina, l’apice della lamina.
La disposizione delle foglie sul fusto è detta fillotassi, è costante per ogni specie e si ripete a intervalli regolari. Nella maggior parte dei casi, le foglie sono poste lungo una spirale; considerando quelle sulla stessa verticale o ortostica, si determina quella che è la disposizione. Le ortostiche si susseguono regolarmente lungo il fusto e conducendo un piano tra due ortostiche e il centro del fusto, individuiamo un angolo al centro che ha un valore caratteristico per specie e generi e dicesi angolo di divergenza.
La spirale lungo la quale si inseriscono le foglie, dicesi spira generatrice.
In base alla fillotassi si distinguono:
Foglie sparse: con spira generatrice lunga e foglie apparentemente sparse.
Foglie alterne: sono una per nodo ma opposte.
Foglie distiche: sono come le alterne ma gli internodi sono molto brevi per cui risultano molto
vicine e riunite in due serie tra loro opposte. (Mimosa, Tasso)
Foglie opposte: sono due per nodo e opposte tra loro.
Foglie tetrastiche: sono come le opposte, ma sovrapposte su nodi alterni.
Foglie decussate: ogni nodo ha una coppia di foglie opposte disposte a 90° rispetto al nodo
contiguo.
Foglie connate: sono come le foglie opposte ma saldate alla base tra loro.
Foglie ternate: ogni nodo porta tre foglie.
Foglie verticillate: ogni nodo porta quattro o più foglie.
Foglie a rosetta: gli internodi sono molto ridotti per cui le foglie sono tutte alla base del fusto.
Foglie unilaterali: le foglie sono tutte dallo stesso lato.
In base a questa caratteristica, le foglie si distinguono in:
erette: formano col fusto un angolo minore di 45°.
patenti: formano col fusto un angolo di 90°.
riflesse: Formano col fusto un angolo maggiore di 90°.
In base a questa caratteristica, le foglie si distinguono in:
sessili: mancano di picciolo.
guainanti: sono attaccate al fusto con la base della foglia o guaina.
picciolate: il picciolo è presente.
amplessicauli: sono sessili e guainanti.
perfogliate: sono guainanti e si ricongiungono dalla parte opposta con i margini.
connate: sono opposte con la parte basale saldata tra loro.
decorrenti: hanno la base allungata in due ali che scendono lungo il caule.
peltate: il picciolo si inserisce al centro del lembo fogliare.
Le nervature rappresentano il sistema conduttore della foglia e sono più evidenti e sporgenti nella pagina inferiore, mentre sono non di rado incavate, nella pagina superiore. Il loro compito è quello di fornire un’impalcatura al lembo fogliare e di distribuire l’acqua e le sostanze assorbite attraverso al radice.
In relazione alle nervature, le foglie si distinguono
in:
anervie: senza nervature (epatiche, muschi)
criptonervie: con nervature non visibili( Aloe)
uninervie: con nervatura unica, non ramificata (Conifere)
pedatinervia: la nervatura si divide in modo dicotomico.
palminervia: le nervature si distaccano tutte dal picciolo.
peltatinervia: le nervature originano dal centro della lamina.
penninervia: da un’unica nervatura centrale si dipartono le
secondarie.
parallelinervia: le nervature decorrono in modo parallelo.
campilodroma: le nervature decorrono seguendo il margine.
Le forme delle foglie sono molto svariate, quindi si ricorderanno le principali:
aghiforme, ensiforme (diritta e appuntita), rotonda, cuoriforme, flabellata (a forma di ventaglio), lineare, lanceolata, ellittica, ovata, spatolata (a racchetta), peltata,ecc.
Intero: il margine è liscio non intaccato.
Ondulato: incisioni ampie poco profonde.
Eroso: incisioni irregolari e numerose.
Sinuoso: incisioni poco profonde.
Seghettato: con dentini inclinati verso l’apice.
Dentato: i dentini hanno i lati uguali.
Crenato: i dentini sono arrotondati.
Lobato: le incisioni dividono la foglia in lobi.
Fido o fesso: con profonde insenature che arrivano circa a metà
del semilembo.
Partito: con insenature oltre la metà del semilembo, ma non
raggiungono la nervatura mediana.
Setto : le insenature raggiungono la nervatura mediana.
La base può essere: arrotondata, troncata, cuneata,
cordata (cuore rovesciato), reniforme (base molto allargata con due grandi
lobi), auricolata (se i lobi sono più piccoli), sagittata (a forma di freccia),
astata (a forma d’alabarda), asimmetrica (le due metà della base sono
diverse).
L’apice può essere: arrotondato, ottuso, acuto,
apiculato (con piccola punta), mucronato (con punta breve e rigida), acuminato
(con una punta), cuspidato (se la punta è molto allungata e sottile),
smarginato (con profonda intaccatura), retuso (con piccola intaccatura),
troncato.
*** *** ***
La foglia è articolata alla base con il fusto,
abbiamo cioè un punto di minore resistenza in cui può staccarsi, è
leggermente ingrossato e dicesi punto di articolazione, talvolta ne
abbiamo diversi nella stessa foglia, in punti diversi, parliamo allora di foglie
composte e la nervatura centrale è detta rachide (Acacia).
La vita della foglia ha una durata variabile che è
legata al tipo di pianta e all’ambiente. Generalmente hanno un anno di vita;
nei paesi con inverno rigido le foglie cadono, all’inizio dell’avversa
stagione. Nel clima mediterraneo è possibile avere piante sempreverdi, quali
l’alloro, l’ulivo, il leccio.
In queste piante, il rinnovamento delle foglie, avviene di continuo, e quelle
vecchie cadono quando le nuove si sono già formate.
Vi sono piante che presentano il fenomeno dell’eterofillia,
ossia la presenza di foglie di forma diversa. Nell’Edera, le foglie basali
hanno cinque punte a differenza delle superiori; nell’Eucalyptus, le foglie
basali sono sessili plagiotrope e le superiori picciolate ortotrope falciformi.
Nello Juniperus sabinae, le foglie adulte sono squamiformi come quelle del
cipresso, mentre le giovani sono normali lesiniformi come quelle delle altre
varietà di ginepro. L’eterofillia può dipendere anche dall’età; in genere
tutte le piante presentano fenomeni di eterofillia, durante il loro sviluppo.
I tipi di foglie che una pianta qualunque presenta
durante l’ontogenesi e che presentano una sorta di eterofillia, sono:
Embriofilli o cotiledoni: sono le prime foglie
che si sviluppano dal seme, diverse dalle foglie
normali, non solo per la forma, ma anche per la funzione. Hanno un alto
contenuto
di sostanze di riserva che saranno utilizzate fino al
raggiungimento dell’autonomia della pianta.
Profilli : sono le prime foglie a comparire dopo i cotiledoni.
Nomofilli : sono le normali foglie adulte, a funzione
clorofilliana.
Ipsofilli o bratte: sono particolari foglie che si
accompagnano al fiore. Quando sono situate lungo
gli assi laterali delle infiorescenze sono denominate bratteole.
Antofilli : sono i costituenti del perianzio, ossia i sepali
del calice e i petali della corolla,
considerati come foglie trasformate.
Sporofilli : presenti nelle felci. Nella pagina inferiore
portano gli sporangi e sono da considerarsi
come fronde, parenti del fusto anche anatomicamente.
Le foglie si possono dividere in due grandi gruppi: le dorso ventrali plagiotrope e le isolaterali ortotrope. Le prime sono le foglie in cui si distingue nettamente il dorso dal ventre; le seconde sono quelle uguali da ambedue le parti, tipo quelle dei giacinti. Per quest’ultime possiamo trovare due tessuti a palizzata, uno per lato della foglia, cioè l’uguaglianza anche anatomica così da avere anche doppia serie di stomi. Le plagiotrope sono le più comuni fra le Dicotiledoni, le ortotrope sono più frequenti fra le Monocotiledoni.
Procedendo dall’esterno verso l’interno, troviamo uno strato d’epidermide sulla pagina superiore e una su quella inferiore, privo di cloroplasti, le due epidermidi si uniscono fra loro ai margini. È uno strato sottile, trasparente e la colorazione della foglia è dovuta agli strati sottostanti. Solo le cellule stomatiche, nello strato inferiore, hanno cloroplasti.
L’epidermide superiore non presenta particolari formazioni, possono essere presenti i peli e le papille, che sono delle piccole sporgenze della lamina.
Nell’epidermide della pagina inferiore, troviamo gli stomi, eccetto che nelle piante acquatiche, dove gli stomi stanno sulla pagina superiore. In genere sono superficiali, ma talvolta infossati in una cripta stomatica, chiusa o no da peli che proteggono da un’eccessiva traspirazione. Altra entità differente, sono gli stomi acquiferi o idatodi. Da essi esce acqua, fenomeno detto di guttazione. Si trovano alla sommità o ai margini della foglia; la guttazione avviene dopo una notte fredda.
Al di sotto troviamo il mesofillo, fatto da parenchima clorofilliano, suddivisibile in due zone, il tessuto a palizzata, e il tessuto spugnoso o lacunoso. Il tessuto a palizzata è formato da cellule più allungate e cilindriche, immediatamente sotto l’epidermide e ad essa perpendicolare. Vi sono spazi intercellulari per favorire gli scambi. Il tessuto lacunoso o spugnoso è formato da cellule isodiametriche, provvisto di cloroplasti in numero vario ma limitato e di colore più chiaro del tessuto a palizzata. È questo tessuto che si collega con le camere stomatiche.
Nel mesofillo troviamo le nervature principali e secondarie. Sono formate da una guaina di tessuto più o meno ispessito che circonda i fasci cribro-legnosi costituenti la nervatura, con il floema nella parte dorsale o inferiore e lo xilema nella parte ventrale o superiore della foglia. La guaina è di tessuto meccanico e a volte di parenchima amilifero. Nelle conifere, possiamo dire di non avere palizzata, ma la parete della cellula s’introflette verso l’interno e in corrispondenza dell’introflessione, si hanno spazi intercellulari.
Nelle foglie si verificano molte modificazioni, indotte per un migliore adattamento della pianta all’ambiente, per una migliore difesa, per una migliore sopravvivenza. Possiamo così avere:
Brattee : si trovano alla base di un singolo fiore o dell’infiorescenza o sotto l’ascella dei peduncoli.
Bratteole : si trovano alla base dei singoli peduncoli di un’infiorescenza.
Cirri: derivano dalla lamina, allorché la rachide
continua circinnata, con funzione prensile o le
rachille (Pisum sativum) che hanno perso la laminazione; oppure è tutta
la foglia a
trasformarsi. In un Lathyrus, tutta la foglia è trasformata in cirro e
sono le stipole che si sono
allargate e hanno sostituito la foglia nelle sue funzioni.
Spine: sono foglie trasformate per ridurre al minimo la
traspirazione. Sono difficili da togliere e
tale operazione lede sempre i tessuti sottostanti.
Ascidi e utricoli: sono propri delle piante
insettivore, in cui esiste del tessuto ghiandolare atto a
digerire l’insetto, con tentacoli di aquixilema d’origine
parenchimatica. Si trova nella
Dionea muscipara, nella Nepenthes. Gli ascidi hanno forma di otri, che
derivano appunto
dalla trasformazione delle foglie, e sono costituiti da: un picciolo, con
funzione di
sostegno, un otre per la cattura degli insetti e una spata fogliare che
serve da coperchio.
Clettrofillo: le foglie presentano il lembo perforato.
Perule: foglie a forma di squame, glutinose a protezione
delle gemme destinate a svernare o
comunque a superare un periodo di sospensione dell’attività
vegetativa. In relazione a ciò,
assumono spesso consistenza, secernendo resine che le impermeabilizzano
(Pioppo,
Ippocastano) o si rivestono di fitte pelurie (Salice).
Catafilli: le troviamo nel bulbo tunicato della cipolla, sono
costituite da parenchima acquifero.
Ocree: stipole trasformate in una specie di manicotto
avvolgente il fusto delle Poligonacee e si
formano per saldatura delle stipole ascellari (si dicono ascellari quando
si saldano in
un’unica lamina).
Fillodi: trasformazione del picciolo che, allargandosi,
assume aspetto fogliare.
Stipole: appendici laminari della base fogliare, spesso
trasformate in spine; nelle graminacee sono
dette ligule
Spata: brattee voluminose che avvolgono l’intera
infiorescenza, lo spadice, nelle Palme e nelle
Araceae e può essere colorata come nell’Anthurium.
Calicetto: involucro che si ha quando le brattee si portano a
contatto diretto col fiore e
contribuiscono alla funzione protettiva.
Galleggianti: tipico esempio è il giacinto d’acqua (Eichornia
crassipes). Il picciolo è molto
rigonfiato per la presenza di parenchima aerifero.
Trasformate: Un esempio l’abbiamo nell’erba pesce (Salvinia
natans), le cui foglie lunghe circa 10
cm., sono costituite da un asse centrale con una doppia serie di piccole
foglie
opposte, la terza foglia è trasformata in filamenti assorbenti.
È la parte dello sporofito con crescita inversa al
caule, si sviluppa quindi sotto terra. Le funzioni principali sono quella di
ancorare la pianta al terreno e la funzione assorbente delle sostanze atte al
nutrimento. Le sostanze nutritive, sono molto diluite nel terreno, per questo è
necessario assorbire anche una grande quantità d’acqua, che sarà poi emessa
con la traspirazione. L’ancoraggio della pianta è molto efficiente e la forza
di penetrazione molto grande. Il fico e il glicine possono penetrare nella
roccia e fenderla. Le radici possono espandersi in superficie ma anche a
notevole profondità. Nelle abetine, ad esempio, abbiamo una radicazione
superficiale in quanto il vento e l’azione dilavante dell’acqua, mettono
allo scoperto le radici, ma l’interazione con radici di piante vicine
assicurano la sopravvivenza delle stesse.
Si può avere, nello stesso individuo, una doppia
radicazione, una profonda e una superficiale, parleremo in questo caso di eterorizia.
La radice è formata dalla continuazione del fusto
sotto terra, non corrisponde però in tutte le sue parti al fusto, solo la parte
centrale di questo si continuerebbe nella radice, mentre la parte periferica si
esaurirebbe nel formare la cuffia pileoriza.
Esaminando la radice distinguiamo: la cuffia
pileoriza o caliptra, a forma di cuneo, attaccata all’apice della
radice, una parte assile senza peli, immediatamente vicina alla pileoriza, una
parte superiore con peli radicali e un punto di contatto col fusto detto colletto.
Nell’embrione, due sono le possibili morfogenesi:
la radice compare come appendice laterale (omorizofite), la radice si prolunga
direttamente (allorizofite). Le omorizofite sono tipiche nelle Pteridophyta,
dove la radice è accessoria all’embrione; nelle Spermatophyta è sempre
allorizofita, questo perché nelle felci, l’embrione non è autotrofo nei
primi momenti di sviluppo, ma è nutrito dal gametofito.
La radice prende origine da un meristema. Si può
avere un meristema intercalare, cioè posto fra due masse, quali la pileoriza e
l’asse della radice, è spostato verso l’apice di una delle due masse, cioè
verso la cima della radice. Lo possiamo definire sub-terminale, in quanto
l’asse della radice è lungo e la cuffia corta. Prolifera da ambedue i lati,
meno in senso centrifugo, più abbondantemente in senso centripeto, produce,
l’asse ad alto numero di cellule e la cuffia di poche cellule.
Quindi, il meristema della radice è sub-terminale,
protetto dalla cuffia, è perciò endogeno, mentre nel fusto era esogeno, ad
apice non protetto. Ciò è in relazione al fatto che, se la radice fosse come
il fusto, spingendosi nel terreno, vedrebbe distruggersi il suo apice; grazie
all’origine endogena, il meristema è protetto e penetra facilmente nel
terreno per mezzo della pileoriza.
Due sono i tipi di radicazione nelle Cormobionta.
Nell’embrione delle Dicotiledoni, troviamo un asse
principale che si ramifica lateralmente, ma l’asse si conserva per lungo tempo
e prevale in lunghezza e grossezza sulle ramificazioni laterali (radici a
fittone). Nelle Monocotiledoni, l’asse si forma inizialmente, ma subito si
atrofizza, ed è proprio in questo punto che nascono nuove radici intorno al
punto di atrofia; sono, se vogliamo, radici avventizie perché nate dopo radici
affastellate o fascicolate.
La cuffia si trova all’apice della radice ed ha
funzione protettiva. È costituita da cellule parenchimatiche, ricche d’amido.
Questo può sembrare strano, dato che dovendo penetrare nel terreno, si potrebbe
pensare alla necessità d’avere cellule molto resistenti, ma la pianta ha
provveduto e se ha cellule tenui, può riformarle con estrema facilità. Questa
formazione avviene in zona centrale, mentre in zona periferica avviene la
progressiva desquamazione. L’apice radicale penetra nel terreno avvolto da
questa guaina desquamante, ma sempre si riforma ed è un enorme vantaggio se si
pensa che sono cellule parenchimatiche, facilmente ricostruibili e che detta
attività dura fino a quando dura il meristema. Si suppone che la presenza di
cellule amilifere, metta la radice in relazione col fusto e vediamo come la
pileoriza sia omologabile alla corteccia, dove pure ci sono cellule amilifere.
Una normale epidermide o rizodermide, liscia senza
peli, separa la cuffia dalla zona pilifera, con centinaia di peli per millimetro
quadro. I peli sono cellule che si estroflettono con all’apice il nucleo, sono
provviste di un citoplasma molto denso, per assorbire, per mezzo del processo
osmotico, i sali del terreno. Data la loro intensa attività, hanno un breve
periodo di vita, e sono sostituiti da altri. Questo vale per tutte le piante, ad
eccezione di quelle desertiche, che hanno una doppia radicazione; col fittone
cercano la falda freatica, con le radici superficiali provvedono ad assorbire la
maggiore quantità di rugiada notturna.
La rizodermide con peli, cessa di funzionare al
momento della desquamazione; la funzione difensiva è ora assunta dall’esodermide,
di origine corticale, perciò ben distinta dalla rizodermide.
Sopra la zona pilifera, troviamo le ramificazioni,
che si suddividono in rami di primo, secondo terzo ordine e così via, e la
struttura sopra descritta per la parte principale, si ripete identica per ogni
ramificazione secondaria.
La lunghezza dell’apparato radicale è molto grande
e così pure lo sviluppo della sua superficie.
Tra le piante, poi, vi è molta differenza riguardo
all’assorbimento; così come esistono piante con poco assorbimento, altre,
come gli Eucalipti assorbono abbondantemente rendendo addirittura impossibile la
vita ad altre piante vicine.
Come primo strato, sopra la pileoriza, troviamo la
rizodermide, presente anche nella zona liscia e nella pilifera. Nella zona
liscia, le cellule sono in definitiva epidermiche, ma essendo prive di cuticola
e di stomi, si preferisce definirle rizodermiche. Nella zona pilifera, sono
presenti i peli radicali, in numero variabile e di dimensioni fra 100 micron e 1
mm. Provvedono ad assorbire acqua, sali minerali e altre sostanze. I peli sono
cellule che si estroflettono e diconsi tricoblasti. Il loro nucleo è in
posizione periferica e il citoplasma è molto denso, l’apice è rotondo.
Rizoderma e peli, hanno in genere, vita breve; la
loro vita è più lunga nelle piante che presentano eterorizia. Col passare del
tempo, il rizoderma desquama e viene sostituito da esoderma che fa parte della
corteccia radicale. Si origina da meristema fondamentale ed ha un parenchima
amilifero di riserva ed è provvisto di tessuto di sostegno, più o meno
indurito, ma sempre elastico. Il parenchima corticale manca naturalmente di
cloroplasti; talvolta può essere stratificato, fino a tre strati, diversi fra
loro, non tanto per la struttura delle cellule, quanto per la loro grandezza. Lo
strato più interessante è il più esterno, detto esoderma, fatto da cellule a
parete ispessita, suberificata internamente, a contatto con la parete primaria.
La suberificazione non è molto spinta, per cui le cellule sono sempre vive. Al
di sotto troviamo endoderma con ispessimenti anche nella parete tangenziale
esterna, per cui in definitiva sono ispessimenti ad U, aperti verso l’esterno.
Quest’ispessimento non interessa tutte le cellule ed è proprio attraverso
quelle che ne sono prive che passano i liquidi verso l’interno, passaggio
assolutamente indispensabile, perché i liquidi, dall’ambiente esterno, devono
in qualche modo arrivare allo xilema.
Si è discusso se questo endoderma appartiene alla
corteccia o alla zona centrale; è meglio considerarlo a sé stante, non omologo
a quello del fusto. All’interno troviamo la stele con un periciclo esterno di
cellule parenchimatiche più o meno simili a quelle del fusto, poi abbiamo la
zona vascolare, ben differenziata da quella del fusto. Non abbiamo, infatti,
fasci cribro-legnosi, ma floema e xilema sono intervallati, cioè la struttura
è alterna, non sovrapposta come nel fusto. Abbiamo qui due sistemi, detti
actinostele piena e actinostele midollata. Quella midollata è più moderna, in
quanto la midollazione è un processo che insorge dopo. Le entità xilematiche
singole sono dette arca, per cui la radice potrà essere tetrarca,
pentarca, ecc. a seconda del numero d’entità xilematiche che presenta.
Infatti, il numero delle arche non è fisso: nelle Conifere sono in genere due;
nelle Dicotiledoni 4-5 nelle monocotiledoni anche 9-11. È un ottimo mezzo
diagnostico.
Queste arche hanno protoxilema esarca, ben visibile,
per cui è facile distinguere una radice da un fusto. Il protoxilema è fatto da
elementi xilematici poco lignificati, con lume piccolo. La posizione del
protoxilema fa pensare che i fasci della radice siano ribaltati rispetto a
quelli del fusto; sorge così il problema del passaggio da una struttura
all’altra. Tra i fasci abbiamo, i raggi midollari, fatti da parenchima
amilifero, al centro, non sempre presente, il midollo.
Come già detto, la radice può ramificarsi più
volte e questo processo può essere di origine endogena, ovvero, il ramo si
forma internamente all’endoderma, in corrispondenza al periciclo e da qui và
verso l’esterno, forando endoderma, corteccia ed uscendo, già provvisto di
pileoriza. È quindi un sistema di ramificazione molto più profondo che nel
fusto. In qualche felce si forma all’interno dell’endoderma e della
corteccia. Quanto descritto appartiene alla struttura primaria il passaggio alla
struttura secondaria avviene come appresso descritto.
In linea di massima abbiamo fasci di floema e xilema
alternati. Insorge ora, un’attività meristematica con la differenziazione di
un cambio (parte tratteggiata), fatto da cellule parenchimatiche che hanno
riacquistato proprietà meristematiche, per cui il cambio è totalmente nuovo.
Sono le cellule sottostanti il floema, che diviene così l’organizzatore di
questo processo, più marcatamente che nel fusto. L’ondata meristematica si
svolge in direzione dei due fasci xilematici contigui, man mano spostandosi
verso l’esterno, a circondare i fasci xilematici stessi. Le cellule del raggio
midollare, ad attività meristematica acquistano così un andamento a serpentina
sinusoidale, lasciano il floema esternamente e circondano lo xilema, che viene
così ad essere interno ad esse.
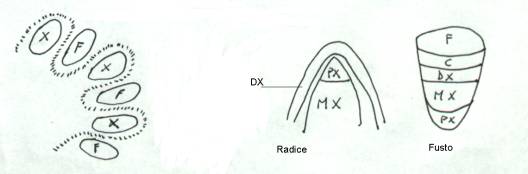
Più tardi, l’anello si distende e finisce per
essere circolare. Mentre nel fusto il protoxilema era endarco e ad esso
seguivano metaxilema, cambio, deuteroxilema, nella radice le condizioni sono
diverse. La figura indica in modo schematico le disposizione del deuteroxilema (DX),
del protoxilema (PX), del metaxilema (MX), del cambio ( C ), del floema (F).
Ancora da ricordare è che il deuteroxilema della
radice non ha alcuna continuità con il cambio primario; nella formazione della
radice il procambio esaurisce tutte le sue forze; nella radice siamo più vicini
allo stato delle monocotiledoni in cui il fascio è chiuso, senza meristema
intercalare.
Mentre il fellogeno del fusto si formava a contatto
dell’epidermide o della corteccia, in definitiva nei primi strati, qui insorge
a livello del periciclo e ciò comporta la desquamazione totale della parte
corticale. Formatosi l’anello cambiale, abbiamo ora una massa xilematica
centrale e una floematica alla periferia, per cui, qualche anno dopo la
formazione completa della struttura secondaria nella radice, si ha piena
corrispondenza con la struttura secondaria del fusto; ma almeno per i primi anni
è ancora possibile il riconoscimento del fusto dalla radice. Infatti, mentre
nel fusto le parti floematiche e xilematiche permangono sullo stesso raggio,
nella radice si trovano su diverse ortostiche.

Due sono le principali teorie, che descrivono il
passaggio dalla struttura del fusto a quella della radice. Occorre tenere
presente che si deve passare da una struttura sovrapposta ad un’alterna, da
protoxilema endarco ad un protoxilema esarco.
Guardando il disegno, il primo cerchio in alto
rappresenta, per tutte e due le teorie, quella che è la struttura del fusto.
Nel secondo cerchio, della I teoria, si vede come le masse si dividano a metà e
migrino nel senso delle frecce. Nel terzo cerchio, della prima teoria, le masse
si sono ricongiunte e il fascio floematico della radice è fatto da due metà
provenienti da fasci distinti del fusto. Bisogna considerare però, la rotazione
nello spazio per cui la massa xilematica ha ruotato anche su se stessa di 180°;
il protoxilema da endarco e divenuto esarco. Le arche restano nella stessa
posizione, solo che sono ora ribaltate. Chi si sposta è il floema; le masse
xilematiche restano sulla stessa verticale, ma ribaltate.
Per la seconda teoria, è la massa floematica a
restare nella stessa posizione, mentre la xilematica, oltre a dividersi in due
metà, si sposta lateralmente e ruota di 180°, per cui da una struttura
sovrapposta si raggiunge l’alterna e si ha pure il cambio di posizione del
protoxilema. Questo processo avviene nell’ipocotile, la parte del fusto
immediatamente sotto all’inserzione dei cotiledoni.
Oggi si ritiene che radice e fusto non siano
perfettamente omologhi, ma che la radice sia solo la parte centrale del fusto.
La parte corticale del fusto darebbe la pileoriza, mentre l’endoderma
corrisponderebbe all’esoderma della radice, corrispondenza suffragata da certe
similitudini, come gli ispessimenti di suberina. Il periciclo del fusto sarebbe
omologo alla corteccia della radice.
Le radici possono presentare delle particolarità e
ne vediamo qualcuna:
Radici avventizie: tipiche sono quelle dell’Hedera che si
originano dal fusto e si attaccano ai muri
o ad altre piante.
Radici coralloidi: sono provviste di tubercoli, dovuti alla
simbiosi con Anabaena cycadearum, una
Cyanophyta
Tubercoli radicali: presenti nelle Leguminose. Contengono il
Ryzobium leguminosarum che fissa
l’azoto atmosferico. È una simbiosi mutualistica in quanto il batterio
permette
alla pianta di assorbire l’azoto fissato, in cambio la pianta fornisce
proteine.
Eterorizia: è il fenomeno per cui una pianta può avere un
doppio apparato radicale.
Piante prive di radici:
la radice nella sua funzione è stata sostituita da funghi endofitici; siamo di
fronte ad una micorriza endotrofica. È presente nelle Ofioglossidae.
Radici respiratorie:
sono importanti nelle zone in cui il terreno ha una struttura tale da essere
asfittico, per cui le radici si modificano opportunamente per respirare.
I rami
secondari escono parzialmente dal terreno, costituendo delle anse dette
pneumatofori, su cui trovansi i pneumatodi. Sono caratteristiche delle
mangrovie tropicali all’estuario dei fiumi, dove il limo rende
asfittico
l’ambiente.
Radici a trampoliere:
sempre nelle mangrovie; il fusto si divide in tante branche che poi
sprofondano nel terreno. Queste piante spesso sono anche vivipare, perché
i
semi germinano attaccati alla pianta madre e restano attaccati fino a
quando
hanno qualche fogliolina e una lunga radice, che si conficca, al momento
del
distacco,
nel terreno e resta fuori per un tratto discreto, così da permettere
alla pianta di nutrirsi e non essere asfissiata.
Radici aeree: tipiche di piante viventi in ambienti molto umidi.
Sono ricoperte di uno strato detto
velamen, che è aquixilema e assorbe acqua. Sono in genere proprie di
piante epifite,
che non toccando terra, difficilmente potrebbero assorbire acqua nel modo
tradizionale.
Radici tuberizzate:
tipiche delle Dalie, contengono sostanze di riserva utilizzate per germinare
Radici tubolari: formano delle lamine larghe 7-10 cm.
talvolta partenti da altezze notevoli. Sono
molto resistenti e su di esse possono impiantarsi piante epifite.
Radici contrattili: in floricoltura, ogni bulbo è piantato ad
una profondità che deve essere in
funzione alla massa; se il bulbo è troppo superficiale non fuoriesce, si
nota
però che si affossa, proprio grazie a radici contrattili che lo tirano
giù.
Austori: tipici del Vischio. È una pianta semiparassita,
che attinge dall’ospite acqua e i sali in essa
dissolti, poi li elabora per suo conto, in quanto essendo verde, ha
capacità di sintesi
clorofilliana. Si attacca grazie ad austori, gli stessi della Cuscuta,
totalmente parassita.
Questi austori hanno una struttura complessa; sono formati da cordoni di
cellule che
s’impiantano in mezzo a cellule parenchimatiche e assorbendo il
nutrimento provocano la
deformazione dei vasi legnosi.
La pianta più antica è l’Aldanophytum, ma la sua
datazione è dubbia, apparterebbe alle Licopsida dell’ultimo Cambriano. Ad
esse appartiene la Baragawanathia, sicuramente del Siluriano. Ora vi sono anche
alcune Rhyniopsida. Nel devoniano siamo già di fronte a più linee filetiche.
Per l’origine, il progenitore più probabile è da ricercarsi fra le
Clorophyta e probabilmente in una Chaetophorales, da cui si sarebbero divise due
linee filetiche conservatrici delle Bryopsida da un lato e delle Stelophytonta
dall’altro, che per successive dicotomie avrebbero originato le altre piante.
Dei gruppi antichi, alcuni si sono estinti, come le Rhyniopsida, altri sono
giunti fino a noi come linee conservatrici. Occorre ricordare che oggi le
Stelophytonta, siano una linea parallela alle Bryophyta, è un gruppo che ha
avuto un notevole potere prospettico, fino a giungere alle moderne piante da
fiore. Per la classificazione, il sistema più moderno è dividerle in due
gruppi e cioè Pteridophyta e Spermatophyta. Alcuni autori invece
adottano una divisione in Lycophyta, Sphaenophyta e Pteridophyta, mettendo in
quest’ultimo gruppo, felci e piante superiori.
Col primo sistema abbiamo due livelli filogenetici ben definiti, sia per l’evoluzione sia per il complesso riproduttivo.