|
|
|
|---|---|
|
Figura 1: |
L'immagine riporta in basso la misura della lunghezza d'onda in metri correlata con le tipologie di radiazioni. In alto si evidenzia lo spettro visibile. È tratta da http://it.wikipedia.org/wiki/Luce |
“In principio Dio creò il cielo e la terra. Ora la terra era informe e deserta e le tenebre ricoprivano l'abisso e lo spirito di Dio aleggiava sulle acque.
Dio disse: <<Sia la luce!>>. E la luce fu.”
Gen 1,1-31
Senza voler parafrasare Isaac Asimov che, nel suo libro “In Principio”, interpreta parola per parola il capitolo della Genesi dal punto di vista storico e scientifico, questa citazione mi permette d'introdurre alcune riflessioni, prima sulla natura del problema e poi sulla sua storia e le possibili relazioni con altri campi del sapere2.
Vedere non è il risultato di soli processi biologici, ma anche psicologici, e interessa vari aspetti dell'esistenza. Per tale ragione, sull'argomento sono intervenuti fisici, fisiologi, esponenti religiosi, filosofi, artisti, neuro-psicologi, linguisti e, da buoni ultimi, chi si occupa di scienze matematiche e d'informatica3. Dal punto di vista storico, tutte le civiltà che si sono succedute nel corso dei secoli, oltre a interrogarsi sull'origine della vita e dell'universo (cosmogonia), si sono chieste quale fosse il rapporto tra l'uomo e il mondo, e in specie si sono interessate alla percezione visiva quale canale di relazione. I filosofi greci, ad esempio, pensavano che il processo visivo fosse dovuto a particelle che uscivano dagli occhi per toccare gli oggetti, come dita che tastano4.
Vedere può apparire intuitivamente semplice, ma una “visione5” più attenta si presenta come fenomeno complesso, la cui funzione è certamente quella di “acquisire una conoscenza del nostro mondo”6, mettendoci in relazione con ciò che ci circonda al fine ultimo di sopravvivere7, reagendo agli eventi che stanno accadendo. È chiaro che per riuscire in questo vi è una stretta interdipendenza con gli altri sensi, in via primaria con il tatto8, che permette di dare certezza alla percezione visiva.
Parlando di sopravvivenza, è d'obbligo richiamare esplicitamente la teoria dell'evoluzione di Charles Darwin, esposta nell'opera “L'origine delle specie”e alle sue conseguenze9. Queste riguardano vari aspetti, che si possono ricondurre, da un lato al mondo scientifico, dall'altro alla sfera sociale. Pur non volendo entrare nel dibattito mai del tutto assopito10 tra favorevoli e contrari, dico subito che nel prosieguo dell’elaborato ritengo tale tesi probabile, e con poche remore11. Darwin stesso, comunque, ammetteva alcuni scogli, come è evidente nel capitolo “Difficoltà della Teoria”12. È singolare che uno di questi riguardasse la formazione di organi perfetti come gli occhi. Le loro capacità, la loro struttura, pongono serie difficoltà a credere che l'evoluzione, attraverso la selezione naturale, abbia potuto raggiungere tale risultato. Ulteriore perplessità era dovuta alle modalità con cui si possono acquisire e modificare gli istinti, quindi non tanto la struttura/forma fisica/biologica dell’animale uomo, ma i suoi comportamenti. Ancora oggi tali problematiche sono, per certi versi, irrisolte e che si ripresentano nel riprodurre artificialmente organi complessi come gli occhi o processi comportamentali-sociali13.
Vedere quindi è essenzialmente un “processo” per l'acquisizione d’informazioni-conoscenza sull'ambiente-mondo che ci circonda (e nel quale siamo immersi) e implica il coinvolgimento di organi biologici e di fattori cognitivi. Nel momento in cui cerchiamo di simulare la vista biologica, da considerarsi come punto di riferimento14 (anche se il parallelismo tra le due presenta delle difficoltà), dovremo valutare non solo le caratteristiche dell'hardware ma soprattutto i requisiti del software, che deve fare riferimento a una conoscenza acquisita15, nonché possedere la capacità di modificarla, tipici temi trattati dall'intelligenza artificiale16.
Per comprendere è quindi necessario conoscere “l'hardware” fornitoci dall'evoluzione biologica; questo potrebbe anche non essere sufficiente, come dichiara David H. Hubel: “All'inizio potremmo pensare che una conoscenza dettagliata delle connessioni dell'occhio con il cervello e dei circuiti all'interno del cervello sia sufficiente a permettere di dedurre i principi generali di funzionamento. Purtroppo questo si verifica solo in minima parte.”17. Ma “Quando abbiamo a che fare con una invenzione dell'uomo, non abbiamo alcun dubbio che il macchinario e le sue parti abbiano funzioni comprensibili. Per capirle è sufficiente leggere le istruzioni relative. Analogamente in biologia, abbiamo sviluppato la fede nel valore funzionale, e in definitiva nella possibilità di comprensione, di strutture che non abbiamo inventato, ma che si sono sviluppate e perfezionate nel corso dei milioni di anni di evoluzione. Il problema del neurobiologo (non il solo, per la verità) è quello di cogliere le correlazioni tra l'ordine e la complessità di una struttura e la sua funzione.”18. Se “l'hardware” non è di semplice comprensione, quando passiamo al “software” il problema si complica ulteriormente, in quanto l'oggetto d'indagine siamo noi stessi e le nostre sensazioni, quindi un campo altamente soggettivo19. Infatti numerose sono le teorie psicologiche che propongono un quadro interpretativo, tra le altre quelle recenti della psicofisica che si legano a quelle della neuropsicologia e della psicobiologia20.
Avendo quindi un'idea di quante e quali possono essere le possibili vie d'approfondimento, iniziamo tenendo presente che la visione umana è fondata sulla percezione della luce, come sottolineano i primi versetti del capitolo biblico della genesi, anche se esistono sistemi alternativi per la costruzione di immagini che si basano su segnali diversi, come l'emissione di raggi laser o onde21 (ad esempio i pipistrelli, o il sonar)... forse gli antichi greci non erano poi così in errore.
La luce è fondamentale non solo per vedere ma anche per vivere. Essa è essenziale per la fotosintesi22 degli organismi vegetali che sono, a loro volta, il primo anello della catena alimentare (non a caso si ritiene i primi organismi viventi furono le alghe grigio-azzurre23). Come potremmo vivere senza la luce? Cerchiamo di comprenderla più da vicino.
Per Newton, è necessario distinguere il mondo fisico dal mondo percepito. Nel primo trattiamo grandezze fisiche che sono oggettivamente misurabili, nel secondo tutto è soggettivo e non misurabile24. La distinzione, fatta per motivi d'analisi, potrebbe generare qualche confusione25 in quanto i due mondi sono collegati: tutti abbiamo esperienza diretta della luce ma definirla a livello fisico non è semplice.
In fisica, la natura della luce è stata oggetto di molte ricerche, tanto che ne costituisce una parte specifica denominata ottica26. Inizialmente, come riporta Richard L. Gregory, si sono affermate due teorie, tra loro rivali: “Isaac Newton riteneva fosse costituita da un flusso di particelle; mentre il fisico olandese Christiaan Huygens sosteneva che gli impulsi luminosi si propagavano attraverso un mezzo impercettibile e omni-pervasivo – l'etere -, concepito sotto forma di sfere elastiche in contatto fra loro: qualunque perturbazione verrebbe trasmessa in tutte le direzioni dalle sfere, accostate le une alle altre, sotto forma di un'onda; onda che costituisce la luce”27.
Possiamo definire la prima teoria corpuscolare e la seconda ondulatoria, ma entrambe non spiegavano completamente il fenomeno. Si è così passati, con James Clerk Maxwell28, alla teoria elettromagnetica, utilizzata oggi nella maggioranza delle applicazioni, e poi alla teoria quantistica di Max Planck29 (ripresa anche da Albert Eistein per la teoria della relatività).
La teoria elettromagnetica migliora quella ondulatoria, interrogandosi sulla natura delle onde30. Lo spazio è ivi definito come campo o spettro elettromagnetico (uno spazio in ogni punto del quale esistono sia azioni elettriche sia azioni magnetiche), e la luce come insieme di onde elettromagnetiche.
Per risolvere alcuni problemi sulla trattazione del corpo nero31, Max Planck propose che lo scambio di energia fra il campo elettromagnetico e la materia potesse avvenire solo tramite pacchetti discreti di energia (quanti) chiamati fotoni. Successivamente Albert Einstein mostrò, studiando l'effetto fotoelettrico, che i fotoni non erano solo un artificio matematico, ma oggetti reali.
Si arriva quindi al concetto dualistico della luce come onda-particella, i quanti “possiedono sia le proprietà caratteristiche dei corpuscoli sia quelle delle onde, tant'è che in molti casi il fotone viene descritto come un “pacchetto” d'onde, composto da un numero tanto maggiore di onde quanto minore è la loro lunghezza.”32
Le onde luminose (formate da un’alternanza di cavi e creste) sono caratterizzate da quattro grandezze fondamentali: l’ampiezza (il massimo spostamento dell’onda in altezza), la lunghezza d’onda (la distanza tra due creste successive), la frequenza (il numero di oscillazioni complete compiute in un secondo) e il periodo (l’intervallo di tempo in cui l’onda compie un’oscillazione completa).
La luce è solo una delle onde elettromagnetiche esistenti. Più precisamente: “Le lunghezze d'onda della luce sono comprese fra 0,38 micrometri33 (3800 Å34, luce blu) e 0,76 micrometri (7600 Å, luce rossa). Per un musicista, il rapporto (1:2) di queste lunghezze d'onda limite (o frequenze) è di un'ottava. L'occhio umano percepisce quindi un'ottava delle oscillazioni elettromagnetiche. La radiazione di 0,5 micrometri corrisponde al colore verde. Una serie di duemila onde di luce verde è lunga un millimetro. Onde più corte di 3800 Å e più lunghe di 900 Å sono radiazioni ultraviolette (UV), mentre le onde più lunghe di 7600 Å (fino a 0,1 mm) sono radiazioni infrarosse (IR)”35. In base alla lunghezza d'onda36 possiamo quindi distinguere sia il colore37 sia il tipo di radiazioni: raggi ultravioletti, raggi X, raggi gamma,luce visibile, raggi infrarossi, microonde, radioonde38. Possiamo anche renderci conto di quanto sia limitata la percezione visiva rispetto alla totalità dello spettro.
|
|
|
|---|---|
|
Figura 1: |
L'immagine riporta in basso la misura della lunghezza d'onda in metri correlata con le tipologie di radiazioni. In alto si evidenzia lo spettro visibile. È tratta da http://it.wikipedia.org/wiki/Luce |
Vi è sempre stato il dubbio se la luce viaggiasse a velocità finita o se raggiungesse istantaneamente qualsiasi regione dello spazio. Nel 1676 Ole Romer39, in base ad alcune osservazioni sull’irregolarità delle eclissi dei satelliti di Giove, concluse che la luce viaggia alla velocità finita di circa 300.000 km/s nel vuoto assoluto (in base alla teoria della relatività di Einstein, è la più alta velocità raggiungibile da un corpo dotato di massa).
Questo significa che, se aggiungiamo al tempo impiegato dai fotoni per raggiungerci i nostri tempi di reazione, vediamo sempre al passato e per poter agire necessitiamo di funzionalità predittive.
La velocità diminuisce nel momento in cui la luce si propaga in un corpo trasparente40 in relazione all'indice di rifrazione41 del corpo stesso (variabile in base alla lunghezza d'onda e alla densità del materiale), permettendo ai prismi di deviare la luce e alle lenti di formare le immagini42. Infatti:“Un oggetto non è visibile se non emette radiazione luminosa. Tranne il caso di oggetti che generano direttamente la luce43, in generale il colore di una superficie dipende dalle caratteristiche della superficie stessa e dalla intensità e direzione della luce che la colpisce.”44. Questo dipende dal fatto che i fotoni viaggiano con traiettorie perfettamente lineari, ma solo finché colpiscono la superficie di un oggetto. In quasi ogni caso la superficie produce un cambiamento radicale nel comportamento del fotone che la colpisce. Sono questi cambiamenti, indotti dalla superficie nel comportamento dei fotoni, che forniscono alla visione le informazioni riguardanti le stesse superfici presenti nell'ambiente. Le sole superfici che non cambiano il comportamento dei fotoni sono quelle completamente trasparenti, quindi letteralmente invisibili (se esistono). Tutte le superfici interagiscono con la luce in modo sufficiente, nella maggior parte delle condizioni ambientali, a essere percepite da un attento osservatore.
Il fotone che incide su una superficie può passarvi attraverso (materiali trasparenti), essere assorbito (corpi oscuri) o essere riflesso (materiali opachi). La luce può passare direttamente attraverso la superficie o essere curvata (rifratta). Di queste interazioni tra fotoni e superfici, la riflessione è la più importante per la visione. Infatti, la luce riflessa è stata cambiata dalla sua interazione con la superficie e contiene, quindi, delle informazioni sulla stessa. Inoltre, la luce riflessa è in seguito disponibile per incidere/colpire la superficie ricettiva dell'occhio di un osservatore, che la può trasmettere così al sistema visivo. Anche la luce non riflessa è importante in quanto permette di comprendere alcuni aspetti della percezione visiva: la luce assorbita per la visione dei colori, la luce passante/trasmessa per la percezione della trasparenza.
Riflettendo un fotone, una superficie ne altera la traiettoria: il fotone rimbalza sulla superficie in una direzione che dipende sia dalla sua provenienza sia dalla struttura microscopica della superficie. Questo significa che le superfici agiscono come fonti di luce secondaria, ossia che la luce non arriva solo direttamente, come nel caso di illuminazione solare senza nuvole o di una lampadina, ma anche in modo riflesso da ogni possibile superficie presente nell'ambiente45.
I fenomeni di riflessione, dispersione e diffrazione della luce sono alla base della nostra percezione di distanze e colori. L’uomo non si limita ad acquisire passivamente i segnali luminosi, ma opera sugli stessi successive valutazioni ed elaborazioni: vedere è un processo attivo.
Prima di analizzare, per quanto possibile, questo processo, propongo alcune riflessioni sulle caratteristiche della luce rispetto ad altri segnali quali il suono, gli odori ecc., nonché sulle interdipendenze tra i vari sensi. La luce si distingue dagli altri segnali per la velocità e la capacità di variazione. La prima ci dà informazioni su oggetti, luoghi/ambienti o avvenimenti lontani da noi, informazioni utili sia per decidere in che direzione muoversi sia per “prepararsi”, decidere “cosa fare”46. La seconda, ci informa sia sulla probabile distanza47 sia, attraverso i colori, sulle probabili qualità degli oggetti stessi (es.: la mela rossa, il cielo scuro, ecc).
Le onde sonore non sono altrettanto veloci48 (es.: vediamo prima il lampo e poi sentiamo il tuono) e non hanno la stessa propagazione. Il segnale sonoro è però facilmente riproducibile (è sufficiente far vibrare una corda, mentre la luce richiede una reazione elettrochimica-magnetica) e semplice da controllare. Pensiamo al linguaggio verbale, al sonar o ai pipistrelli che grazie al fenomeno dell'eco possono localizzare gli oggetti/ostacoli presenti nell'ambiente (eco-localizzazione). Al limite c'è anche una differenza d'approccio: con la luce noi ci limitiamo a recepire delle onde/particelle che comunque andrebbero a colpire l'oggetto in quanto emesse da una sorgente per noi generalmente incontrollabile (es.: il solare), comunque di grandezza infinitamente piccola, quindi non modifichiamo l'oggetto (l'eccezione potrebbe essere rappresentata da organismi fotosensibili, ma anche i sali d'argento usati in fotografia), insomma non siamo invasivi e potremmo essere anonimi. Poi, il suono comporta il movimento di materia, di atomi se non di molecole (es.: aria, acqua), quindi anche delle sollecitazioni fisiche sugli oggetti (ad esempio mi è capitato di sentir vibrare i vetri delle finestre in occasione di lancio di fuochi d'artificio per feste paesane) perciò se utilizziamo il suono siamo più invasivi, la fonte del suono è quasi sempre identificabile, se siamo noi, probabilmente non siamo anonimi.
Anche l'olfatto ci permette di avere informazioni a distanza, limitate però in portata e precisione (metri, decine di metri, chilometri?), in quanto l'odore non si propaga in modo uniforme. L’olfatto è d’altro canto molto più attendibile per quanto riguarda la composizione chimica delle sostanze (es.: gpl, metano, carne avariata, pane, biscotti appena sfornati, ecc).
Con il tatto acquisiamo certezze: possiamo stabilire ciò che è solido, ciò è ruvido, ecc. Ma se abbiamo certezze significa anche che siamo coinvolti da ciò che ci circonda, nel bene, se questo ci fornisce delle possibilità (nutrirci, ecc), e nel male, (come toccare un metallo incandescente).
Infine il gusto, che come l'odorato ci dà informazioni chimiche sulla sostanza che abbiamo già assunto al nostro interno, dandoci un'ultima possibilità di espellerla prima che provochi danni.
Penso sia chiaro come la nostra sopravvivenza sia strettamente legata alla conoscenza del mondo/ambiente/universo che ci circonda, quindi alla nostra capacità di reperire informazioni il più possibile affidabili, ossia avere una percezione veridica. Interessante quanto afferma Stephen Palmer: “Questo è quasi sempre il caso della visione, ed è per questo, probabilmente, che la prendiamo quasi sempre per sicura. Sembra quasi una finestra perfettamente trasparente sulla realtà. Ma è veramente così? [...] Io argomenterò che la visione non è una finestra trasparente sulla realtà, ma una costruzione attiva, un significativo modello dell'ambiente che permette al percettore di predire che cosa succederà nel futuro così da poter effettuare azioni appropriate, aumentando le possibilità di sopravvivere.”49.
Se la luce è il mezzo, il canale, per la diffusione dell'informazione visiva, è necessario disporre anche di un apparato recettore in grado di captarne i segnali. È utile a tal fine un'analisi che confronti i vari sistemi sia nel corso dell'evoluzione, a livello diacronico, sia al momento attuale nei vari organismi, a livello sincronico.
Come afferma Richard L. Gregory è necessario, in aggiunta, considerare i rapporti tra l'apparato recettore e quello che si occuperà di elaborare l'informazione: “A cosa potrebbe mai servire un cristallino fatto a metà, o anche un cristallino in grado di focalizzare un'immagine, se non vi fosse poi un cervello capace di servirsene? E come potrebbe d'altronde essersi formato un cervello con tali capacità prima che vi fosse l'occhio in grado di fornirgli informazioni visive? In ambito evolutivo non può esservi alcuna programmazione, alcun tipo di previsione, alcun controllo sperimentale per eliminare quanto può essere inefficace. Occhio e cervello si sono evoluti attraverso un processo lento e casuale di tentativi ed errori. Per ripercorrere le tappe dello stesso dobbiamo cercare di rintracciare i possibili vantaggi incrementali di ciascuno stadio, seppure ammettendo che qualcosa di vantaggioso per un verso possa poi rivelarsi utile in un campo totalmente differente.”50.
Non solo alcuni minerali sono sensibili alla luce: anche a livello biochimico, quindi nei vegetali e negli animali, avvengano delle reazioni basate sulla luce51. Ad esempio è facile osservare come alcune piante dotate di mobilità attiva seguano lo spostamento della luce.
L'analisi della probabile evoluzione ce la fornisce ancora Gregory con alcuni esempi: “La reazione alla luce si verifica perfino in esseri unicellulari, e nelle forme animali più evolute troviamo cellule specializzate che funzionano come fotorecettori sensibili al movimento. Queste cellule possono essere sparse su tutta la pelle (come nel lombrico) oppure raggruppate, spesso lungo una piega cutanea o in una depressione della superficie corporea, secondo un adattamento che prelude al vero e proprio occhio capace di formare le immagini.
Sembra probabile che gli elementi fotorecettori si siano collocati in questi recessi per trovare protezione dalla luce abbagliante, che riduceva la loro capacità di individuare ombre in movimento, segnali dell'avvicinarsi di un pericolo. In queste loro sedi primitive i recettori erano sottoposti al rischio di venire occlusi da polveri e piccoli corpi estranei, che sovrapponendosi a essi potevano precludere la visione della luce. Per ovviare a tale inconveniente si formò allora, per effetto di mutamenti fortuiti, una membrana di protezione; la quale, in seguito, divenne più spessa al centro e si trasformò in una vera e propria lente (il cristallino). In un primo tempo il cristallino serviva soltanto per potenziare la luminosità, ma in seguito venne a formare immagini effettive. Un esempio di occhio primitivo si può ancora osservare in un mollusco marino, la patella52; e un altro mollusco, il Nautilus53, è fornito di un occhio ancor più primitivo, privo di cristallino ma provvisto di un piccolo foro che serve a formare le immagini. L'interno dell'occhio del Nautilus è lavato dal mare in cui esso vive, mentre gli occhi più evoluti, simili al nostro, sono pieni di un liquido appositamente prodotto (l'umor acqueo) che sostituisce l'acqua di mare; e le lacrime umane, con il loro sapore salato, sono una sorta di riproduzione degli oceani primordiali che bagnavano i primi occhi.”54
Gregory prosegue analizzando il funzionamento dell'occhio degli invertebrati55 che ha una struttura più complessa dell'occhio umano poiché è formato da molte lenti con un piccolo gruppo di recettori, circa uno per ogni singolo cristallino. Esso ha delle prestazioni migliori di quello dell'uomo specie nella rilevazione del movimento: a riprova di ciò si pensi alla libellula che cattura le prede in volo, o alla capacità di recepire i raggi ultravioletti, come nel caso delle api. Interessanti, negli occhi composti, sono i meccanismi che permettono l'adattamento alla luce e alla penombra, o il funzionamento della lente cilindrica, che per le sue caratteristiche richiama i moderni strumenti utilizzati in endoscopia medica o le fibre ottiche56.
Gregory, passando attraverso l'analisi del sistema visivo della Copilia (un microorganismo acquatico57), arriva ad ipotizzare “che l'elaborazione visiva sia derivata dall'elaborazione della sensazione tattile, caratterizzata da una utilità immediata ed essenzialmente più semplice. Può essere suggestivo pensare che vi siano due tipi di tatto: una ricezione tattile attiva ottenuta muovendo le ciglia, o le dita, e una ricezione passiva legata alla percezione delle forme. In effetti, si può scorrere lentamente con le dita sulla superficie degli oggetti, oppure sperimentare direttamente la forma di piccoli oggetti, nella loro unitarietà, portandoli a contatto con un'area estesa della pelle.”58. Così facendo egli supera l'impasse dovuta alle possibili differenze evolutive tra occhio e cervello.
Non solo, l'originale meccanismo visivo a scansione utilizzato dalla Copilia potrebbe fungere da esempio per strumenti dediti a particolari applicazioni, e permettere di introdurre una riflessione sull'evoluzione epistemologica della scienza. Infatti, quando nel 1891, Selig Exner osservò che il sistema ricettivo della Copilia e la lente cilindrica ad esso unita compivano "dei continui movimenti attivi", egli non riuscì a darne una spiegazione. Fu con l'avvento della televisione che si ebbero le nozioni necessarie alla comprensione. La scienza, e noi con essa, ha quindi bisogno di “modelli di riferimento” per comprendere, avere una conoscenza più completa o avvicinarsi alla verità59.
La brevissima panoramica sulla possibile evoluzione dei sistemi ottici e l'ipotesi che la vista discenda dal tatto dovrebbero trasmettere sia l'idea della complessità dell'argomento, sia il fatto che la natura ha risolto con “dispositivi” diversi il problema della percezione visiva. Nel riprodurla dovremo quindi valutare qual è l'obbiettivo e quali le possibili criticità. Con questo fine, cerchiamo di comprendere le peculiarità dell'occhio umano, quale modello di riferimento, analizzandone le funzionalità e la struttura.
|
|
|
||
|
Figura 2 |
Sezione orizzontale obliqua dell'occhio http://it.wikipedia.org/wiki/Immagine:Occhio_umano_spaccato.png 13 settembre 2006 |
Figura 3 |
http://it.wikipedia.org/wiki/Immagine:Occhio_umano.png 13 settembre 2006 |
Il sistema ottico può essere considerato come un meccanismo che converte/trasforma gli stimoli/energia luminosa che si imprimono nella retina in un flusso in uscita simile a un segnale elettrico. Come si può osservare dalla fig. 2, la luce entra attraverso la cornea, passa in sequenza la camera anteriore, la pupilla (apertura nell'iride), il cristallino, e il corpo vitreo (o humor vitreo), prima di imprimersi sullo strato di fotorecettori che costituiscono il retro della retina. Questa è responsabile della reale trasformazione da energia luminosa in segnali nervosi nella forma di una sequenza d'impulsi a frequenza modulata61.
Nonostante la sua forma globulare avente un diametro di circa 25 mm, l'occhio umano è funzionalmente simile a una telecamera con un campo visivo che copre un'area di circa 160º in ampiezza per 135º d'altezza. Come ogni altro sistema ottico, soffre di vari tipi di distorsioni geometriche e cromatiche62. Si tenga presente infatti che solo il 50 per cento della luce che attraversa la cornea arriva alla retina. Il sistema ottico quindi proietta un’immagine riconoscibile ma molto imperfetta sulle cellule recettrici63, tuttavia in quantità e qualità sufficiente a fornire al sistema visivo informazioni eccellenti riguardanti l'ambiente64.
Oltre alla struttura interna è opportuno valutare il posizionamento degli occhi e il loro numero. Essi sono collocati sulla linea immaginaria che divide orizzontalmente a metà la testa, all'interno di due fori quasi semisferici del cranio, denominati orbite oculari, che li tengono saldamente in posizione e permettono, attraverso la rotazione, di dirigerli come si desidera o necessita. Ogni occhio si muove grazie a sei piccoli ma possenti muscoli denominati extraoculari, controllati da specifiche aree del cervello. Il loro numero è in ragione del loro funzionamento: sono organizzati in tre coppie, e i muscoli di ogni coppia sono antagonisti, così da determinare movimenti in uno dei tre piani ortogonali (perpendicolari) dello spazio. I movimenti sono necessari per sondare diverse regioni del campo visivo senza dover muovere l'intero capo e per la messa a fuoco di oggetti posti a distanze diverse. Per i due occhi, il compito di seguire un oggetto deve essere svolto con una precisione dell'ordine dei minuti di arco65, altrimenti vedremmo doppio66. Palpebre e ciglia proteggono l'occhio, le lacrime lo tengono umido e pulito.
Alcuni di questi aspetti sono veri anche per occhi non umani. Tuttavia alcune specie, come i piccioni e i gufi, non possono muovere gli occhi nelle orbite e sono costretti a spostare l'intero capo. Questo limita la velocità con cui essi percepiscono gli eventi nell'ambiente, semplicemente per il fatto che la testa è più pesante e difficile da spostare degli occhi. In aggiunta la posizione differisce in modo notevole nelle varie specie. Nell'uomo gli occhi sono entrambi posizionati nella parte frontale del capo, mentre in molti animali si trovano più vicini ai lati. Il posizionamento frontale fornisce un campo visivo con un'ampia area di sovrapposizione dei due occhi, limitandone però la copertura totale. Il beneficio della sovrapposizione del campo visivo (detta anche visione binoculare o stereovisione) consiste nel permettere la percezione della profondità. La capacità di stimare la distanza in modo attendibile è un vantaggio evolutivo per i predatori, che hanno bisogno dell'informazione per attaccare: è per questo che gli occhi dei cacciatori tendono a essere posizionati frontalmente con sovrapposizione dei campi. All'opposto, campi visivi panoramici sono vantaggiosi per le possibili prede per monitorare in modo più esteso possibile l'ambiente che le circonda al fine di individuare possibili pericoli, quindi gli occhi sono posizionati lateralmente. Altri animali hanno gli occhi posizionati frontalmente o lateralmente per ragioni diverse, ma tutte riflettono lo stesso bilanciamento tra un‘accurata percezione della profondità e copertura dell'ambiente67.
A questo punto, date alcune informazioni sulla composizione della luce e sulla struttura dell'occhio, è opportuno chiedersi, prima di vedere più in dettaglio alcuni componenti dello stesso e quanto (relativamente poco) si conosce del cervello, come si formano le immagini ottiche.
Platone (427-347 a.C.), filosofo greco, credeva che un “fuoco interno” facesse nascere dei raggi che partivano dagli occhi verso gli oggetti percepiti. Questa teoria, denominata emanazione o emissiva, venne ripresa da Euclide (365-275 a.C.), probabilmente discepolo di Platone, e inserita nella sua opera “Ottica”. Epicuro (341-270 a.C.) la rigettava credendo, invece, che piccole repliche degli oggetti fossero, in qualche modo, rapidamente trasmesse nella mente attraverso gli occhi. Galeno (130-200 d.C.) elaborò l'idea con dettagli fisiologici, proponendo che dopo l'emanazione dei raggi questi interagissero con gli oggetti e ritornassero agli occhi. Egli credeva che nel cristallino i raggi, interagissero poi con uno “spirito visivo” che va e ritorna dal cervello agli occhi portando con sé le repliche degli oggetti percepiti68. Fu Alhazen (965-1040 d.C.), filosofo arabo, che intuì come l'occhio funzionasse in modo simile a una camera con un foro in una parete69 che permetteva la formazione, nella parete opposta a quella forata, di immagini interne, anche se capovolte, delle figure di oggetti esterni alla camera. Egli progettò così la camera oscura che fu successivamente perfezionata da Giovan Battista Della Porta (1535-1615 d.C.), che inserì nel foro una lente focalizzante, per ottenere immagini più brillanti e riuscire a individuare forme e colori70. Fu infine Johannes Kepler (Keplero 1571-1630 d.C.) che formulò la moderna teoria fisiologica dell'ottica con un'approssimazione accettabile. Teoria successivamente sperimentata da padre Cristoph Scheiner (1573-1650 d.C.) nel 1625 e adottata da René Descartes (Cartesio 1595-1650 d.C.)71.
|
|
|
||
|
Figura 4 |
Descrizione di camera oscura di Giovanni della Porta (1544). http://puertadelarte.com/arteporta/talleres.htm 7 ottobre 2006 |
Figura 5 |
Camera oscura portatile tipo reflex. http://www.writers.it/art/articoli/scheletri/intro.htm 7 ottobre 2006
|
La formazione delle immagini ottiche è quindi la trasformazione di un mondo che ha tre dimensioni spaziali in rappresentazioni bidimensionali. Gli oggetti sono immersi in spazzi tridimensionali illuminati dalla luce, che viene riflessa dalle loro superfici nell'occhio dell'osservatore lungo linee rette. I fotoni entrano nell'occhio formando un'immagine bidimensionale capovolta sul fondo. L'oggetto esterno è spesso riferito come stimolo distale (significa lontano dall'osservatore), e la sua proiezione sul retro della retina come stimolo prossimo (vicino all'osservatore). La grandezza dell'immagine dell'oggetto nell'occhio è solitamente definita dal suo angolo visivo: il numero di gradi sottesi dall'immagine dai suoi estremi fino al punto focale dell'occhio. Quest'angolo misura la dimensione spaziale dello stimolo prossimo e non quello distale. Lo stesso oggetto esterno sottenderà un angolo inferiore quando è lontano dall'osservatore e uno maggiore quando gli sarà vicino. La relazione tra dimensione e distanza dell'oggetto e dimensione dell'immagine è importante per comprendere come percepiamo la grandezza e il posizionamento degli oggetti72.
Forse il fatto più importante, per quanto riguarda il processo di formazione delle immagini e la comprensione del processo visivo, è che l'immagine sul fondo dell'occhio ha solo due dimensioni spaziali al posto di tre, cioè è simile alla proiezione su una superficie curva. Questo significa che vitali informazioni riguardanti lo spazio si perdono passando dal mondo reale tridimensionale (in seguito 3-D) alla sua rappresentazione oculare bidimensionale (in seguito 2-D). La grandezza persa è la profondità: la distanza che va dal punto focale d'osservazione alla superficie ambientale che riflette la luce. Per percepire il mondo in 3-D la profondità deve essere in qualche modo recuperata dalle informazioni fornite dall'immagine ottica bidimensionale73.
Il processo di trasformazione da 3-D a 2-D ubbidisce a determinate leggi fisiche e può quindi essere analizzato matematicamente. Nel caso specifico si usano le proiezioni geometriche: gli studi di come uno spazio a dimensioni superiori è mappato in uno a dimensioni inferiori. La proiezione geometrica può determinare esattamente, per una data scena 3-D, dove ogni punto di questa sarà proiettato in un piano 2-D creando la relativa immagine, e quali saranno le proprietà invariabili delle possibili immagini in varie proiezioni. Nella visione dinamica, la proiezione, della scena considerata, parte dalle quattro dimensioni strutturali dello spazio-tempo, passa per lo spazio tridimensionale del flusso ottico che si stende nel tempo e arriva alle superfici bidimensionali nel fondo dell'occhio.
La proiezione geometrica sembra perciò essere lo strumento matematico ideale per comprendere la formazione delle immagini. Il problema è che da sola non può riprodurre la complessità dei fenomeni ottici in quanto non contiene le strutture appropriate per la modellazione della riflessione della luce, dell'assorbimento o della rifrazione. In un mondo saturo di superfici opache, ad esempio, solo la luce riflessa da superfici vicine raggiunge l'occhio. I fotoni provenienti da punti lontani saranno o assorbiti o riflessi dalle superfici più vicine, senza arrivare, perciò, agli occhi.
Per superare questi problemi si ritorna al modello della camera oscura, quindi alle osservazioni fatte da Alhazen. Poiché i fotoni viaggiano in linee rette, la luce che cade su ogni punto del piano dell'immagine della camera oscura vi giunge riflessa (o emessa) da un punto specifico dell'ambiente. Questo punto sta sul raggio che parte dal punto sul piano dell'immagine esterno e che attraversa il foro della camera. Il tutto è facilmente comprensibile nelle figure 4 e 5, che sono prime rudimentali riproduzioni di fenomeni osservabili, ma sopratutto in figura 6, che rappresenta lo schema di funzionamento di una macchina fotografica e/o dell'occhio74. Il punto “posizione” è all'interno della camera oscura, quindi è il piano su cui cade/incide il raggio di luce, il punto “look_at” o “mira” è un punto nello spazio ambientale, il “piano dell'immagine” è la parete con il foro o l'eventuale lente. Per questi punti passa la linea retta percorsa dai fotoni.
Questa situazione dà la proiezione prospettica (o polare) della geometria di base: il processo di formazione dell'immagine dove la luce converge in un unico punto focale (o polo).
|
|
|
|
Figura 6 |
Proiezione prospettica: in figura la proiezione simula la classica macchina fotografica, e riproduce il normale effetto di prospettiva. http://www.liberliber.it/biblioteca/p/pov_ray_team/manuale_pov_ray_v3_01/html/741.htm 07 ottobre 2006 |
Buone foto realizzate con la camera oscura non sono così semplici da ottenere come la presente descrizione può far sembrare. Per avere un’immagine chiara e vivace, è necessario che il foro sia molto piccolo; circa 0,4 mm di diametro sono l'ideale. A causa di questa ridotta apertura, solo pochissima luce incide sulla parete interna. Se il foro viene allargato, facendo passare più luce, l'immagine diventa confusa poiché tutta la luce non passa più per un singolo punto ma attraverso più punti. Questo problema viene superato ponendo una lente convessa trasparente (un obbiettivo) sul foro, così da curvare i raggi di luce entrate in un punto (detto punto focale) dietro alla stessa lente ma leggermente distanziato. Perciò l'obbiettivo fornisce un “foro virtuale” con un suo punto focale che rende l'immagine proiettata sul fondo nuovamente definita e chiara, con il vantaggio, rispetto alla camera oscura di partenza, di essere più brillante in quanto entra più luce. Nell'occhio umano sia cornea che cristallino cooperano per la messa a fuoco.
|
|
|
||
|
Figura 7 |
Schema di proiezione ortogonale: la proiezione ortogonale di una qualunque figura, piana o solida, è la proiezione ortogonale di tutti i punti che la compongono. http://www.aldodirosa.it/news_docenti/html/modules.php?name=Proiezioni1 07 ottobre 2006 |
Figura 8 |
Esempio di proiezione ortogonale: il considerare i raggi luminosi provenienti da un punto improprio significa che questi sono paralleli fra di loro e la proiezione, su di un piano, del contorno apparente dell'oggetto illuminato segue una proiezione parallela (proiezione ortogonale). http://digilander.libero.it/cartesolari/ombra.html 07 ottobre 2006 |
Come si può ben intuire le elaborazioni matematiche relative alla proiezione prospettica sono complesse. Al fine di semplificarle spesso usata una “prospettiva debole” denominata proiezione ortografica o parallela per fornire un modello geometrico della formazione dell'immagine. In questo caso si considera l'immagine come formata da raggi di luce che viaggiano paralleli gli uni agli altri e perpendicolari al piano dell'immagine, piuttosto che raggi che convergono sull'obbiettivo o sul foro della camera oscura.
La proiezione ortogonale permette di semplificare l'elaborazione matematica in quanto ignora la dimensione di profondità, le distanze dall'immagine agli oggetti, del mondo, mentre tutte le informazioni spaziali sul piano perpendicolare alla direzione visiva sono preservate senza cambiamenti. Questo significa che, quando la distanza che va dall'immagine all'oggetto è sufficientemente grande, rispetto alla profondità dello stesso, la proiezione ortografica è una buona approssimazione della proiezione prospettica. Da vicino, invece, le differenze sono significative. Quindi, la proiezione ortografica può essere concepita come un caso limite della proiezione prospettica, dove la distanza tra l'oggetto e il punto focale è infinita. La differenza è che nella seconda a una distanza infinita l'oggetto è un punto, nella prima è un immagine estesa nello spazio75.
La visione è quindi il problema inverso alla formazione delle immagini: come riottenere da immagini ottiche di una scena la conoscenza degli oggetti che la formano. Sebbene sembri facile invertire il processo, esso risulta alquanto difficile, se non impossibile. Se da uno spazio tridimensionale si può ricavare una sola immagine bidimensionale, non è vero il contrario, ossia, ogni punto dell'immagine può essere mappato in un infinito numero di punti nell'ambiente. Si sa che è possibile solo in quanto il sistema visivo umano vi riesce con accuratezza in molte circostanze. Sul come vi riesca, vi sono numerose teorie, la più nota delle quali afferma che tale sistema elabori numerose assunzioni altamente probabili circa la natura dell'ambiente e le condizioni in cui è visto. I vincoli posti da queste assunzioni permettono di risolvere il “problema inverso” nella maggior parte dei casi. La visione è per questo un processo euristico, dove si inferiscono le condizioni più probabili dell'ambiente che ha prodotto l'immagine. Il processo è euristico in quanto usa regole empiriche di inferenza – basate su assunzioni addizionali – che non sono sempre valide e quindi, qualche volta, possono portare a conclusioni errate. Poiché sono elaborate nel cervello le vedremo in seguito, ora esaminiamo nel dettaglio i componenti dell'occhio seguendo il percorso fatto dai fotoni.
La cornea (che è coperta da una pellicola di lacrime) è la superficie frontale dell'occhio che curva la luce per formare l'immagine. È trasparente ed è sorretta da uno strato opaco di membrana fibrosa denominata “sclera”76. Essa si caratterizza per la mancanza di vasi sanguigni e per questo ricava il suo nutrimento dall'umore acqueo, questo la rende praticamente indipendente dal resto dell'organismo (per questa ragione i trapianti di cornea sono sicuri: gli anticorpi non possono raggiungerla e distruggerla). L'umore acqueo è completamente rinnovato ogni quattro ore circa, in quanto viene continuamente secreto e assorbito; alcune volte, in questo liquido, sono presenti delle impurità che compaiono davanti agli occhi e, poiché proiettano la loro ombra sulla retina, vengono viste come macchie vaganti nello spazio77. Il percorso aria-cornea-umore acqueo è responsabile di approssimativamente due-terzi della capacità ottica dell'occhio (42 diottrie contro le possibili da 57 a 62 totali).
Dopo aver attraversato la cornea e l'umore acqueo la luce passa attraverso la pupilla, un diaframma o apertura al centro dell'iride, che è, appunto, un muscolo a forma anulare. Il pigmento78 presente nell'iride assume una vasta gamma di colori. Non a caso, iride significa arcobaleno, in greco.
La funzione dell'iride è di contrarsi, restringendo il foro pupillare, sia quando la luce è intensa, per ridurre l'apertura della lente, sia quando gli occhi convergono, per vedere oggetti vicini. Ciò aumenta la profondità di campo che consente di mettere a fuoco distintamente, il processo è analogo a quello della riduzione del diaframma in una macchina fotografica79. Le variazioni del diametro del diaframma sono ottenute con la contrazione di due fasci di muscoli lisci, lo sfintere e il dilatatore pupillare. Lo sfintere è responsabile della costrizione e ha un moto circolare parallelo all'iride; il dilatatore, con fibre radiali disposte come i raggi di una ruota, apre la pupilla. In cooperazione questi muscoli possono posizionarsi in 16 diverse aperture variando il diametro da 2mm a 8mm. Per massimizzare la messa a fuoco il meccanismo di controllo tende a tenere l'apertura più stretta possibile. L'operazione richiede circa 10s per l'intero diametro. L'attivazione del controllo è estremamente complessa e dovuta a numerosi fattori, ma principalmente alla quantità di luce incidente e allo stato emotivo dell'osservatore. In verità, una recente indagine ha fornito una lista di 23 diverse cause di variazione, tra cui le preferenze sessuali, le attitudini politiche, la fatica,e la lunghezza d'onda di colori e suoni, nonché alcuni disordini neurologici.
Riassumendo, variare la dimensione della pupilla ha tre obbiettivi. Il primo, come sopra esposto, consiste nel controllare la quantità di luce che s'imprime sulla retina. Il secondo, conosciuto come risposta alla vicinanza, tende a mettere a fuoco gli oggetti vicini, quindi adeguarsi alla profondità di campo. Il terzo, importante in condizioni di luce brillante, è la riduzione dell'apertura per evitare distorsioni dell'immagine. Sembra che solo i primi due siano quantitativamente controllabili da stimoli esterni80.
Oltre la pupilla, la luce passa attraverso il cristallino, che fornisce il terzo rimanente del potere diottrico. Il suo compito principale è di compiere gli aggiustamenti necessari per mettere a fuoco oggetti a distanze diverse. Nell'uomo la lente cristallina è formata da un insieme di strati sottili, tra loro sovrapposti, ed è fissata da una membrana, la zonula, che ne regola, anche, lo stato di tensione81. Il funzionamento è simile a quello della messa a fuoco di una macchina fotografica: si varia la distanza tra la lente e la pellicola; nell'uomo, invece, cambia la forma del cristallino, mettendo in tensione o rilasciando i tendini che lo fissano alla periferia, così può diventare più sferico per vedere gli oggetti vicini e più appiattito per vedere quelli lontani. Queste variazioni di forma sono determinate da un insieme di muscoli radiali chiamati ciliari. Il processo avviene in tempi rapidi, circa 0,4s (dopo i quarantacinque anni, il cristallino si irrigidisce e viene meno la capacità di messa a fuoco: per superare questo problema Benjamin Franklin inventò gli occhiali bifocali). Lo stimolo necessario a far contrarre i muscoli ciliari, e modificare la forma del cristallino, è dato dall'input visivo, questo, a sua volta, è collegato al riflesso che controlla la convergenza degli occhi82.
L'umore vitreo è un liquido gelatinoso responsabile del mantenimento della forma del globo oculare. La luce passa attraverso il liquido alla retina, che a sua volta si occupa della conversione elettro-ottica delle forme di luce che incidono sui fotorecettori della retina 83.
Il termine retina etimologicamente discende da "rete", o "tunica a forma di ragnatela", ed è dovuto al fatto della presenza di un fitto intreccio di vasi sanguigni nella membrana retinica84.
Prima di esaminarne la struttura ricordo, come accennato nel paragrafo “Formazione delle immagini”, che fu Keplero, nel 1604, a rendersi conto che la retina è il primo stadio della percezione visiva: essa infatti costituisce lo schermo su cui vengono proiettate le immagini provenienti dalla lente cristallina. L'ipotesi è stata in seguito controllata sperimentalmente nel 1625 da Scheiner che, prima asportò la parte posteriore dell'occhio di un bue delle membrane più esterne (la sclerotica e la coroide), quindi, osservò una piccola immagine capovolta sulla retina lasciata allo scoperto. La retina appariva simile a una pellicola diafana (trasparente, esile). Successivamente, Cartesio ripeté l'esperimento, descrivendolo nella sua Diottrica del 1637, e vi aggiunse delle chiare schematizzazioni. È sorprendente che la scienza sia giunta a questa scoperta relativamente tardi nella sua storia, e ancor più strano è che non sia stato apprezzato fin da subito l’ampio spettro delle sue implicazioni. Oggi vi è ancora chi tenta di conservare l'idea secondo cui la percezione è direttamente in contatto con il mondo degli oggetti, ignorando la presenza dello schermo ottico della retina e la complessità fisiologica del processo di elaborazione dell'immagine necessario a creare le percezioni. Il sapersi isolati dal mondo esterno può, probabilmente, essere fonte di timore e, portare ad illusioni ottiche85.
La retina è una parte del cervello, ma si è separata da questo nel corso dell'evoluzione; tuttavia vi è ancora un fascio di fibre che funge da connessione tra le parti: il nervo ottico. La retina ha la forma di una lamina, come altre strutture del sistema nervoso centrale. Il suo spessore è di circa un quarto di millimetro. È costituita da cinque strati: tre strati di neuroni, separati da due strati che contengono le sinapsi fatte dagli assoni e dai dendriti86, come visibile in figura 10.
|
|
|
||
|
Figura 9
|
Schema di trasmissione di segnali in neuroni e del neurone, di proprietà di P. Forster. http://it.wikipedia.org/wiki/Neurone 9 ottobre 2006. |
Figura 10 |
Schema dei livelli neurali nella retina, tratto da Henry Gray (1825–1861). Anatomy of the Human Body. 1918. http://www.bartleby.com/107/illus882.html 10 ottobre 2006 |
Prima di esaminarli è opportuno comprendere come sia fatto e come funzioni l'elemento base del cervello e della retina: il neurone. Esso è una cellula specializzata che, grazie ad una complessa serie di reazioni biochimiche interne, unisce l'attività di altri neuroni (come input) ad essa collegati e ne diffonde/propaga (in output) il risultato ad altri in successione87. Il neurone può essere suddiviso in cinque parti:
Dendriti: sono delle ramificazioni che partono dal corpo della cellula nervosa. Sono specializzati nel rispondere ai segnali provenienti da altri neuroni o dall'ambiente esterno. La loro forma ramificatali, rendendo disponibile un'ampia superficie, li rende adatti alla ricezione dei segnali. I dendriti dei neuroni sensoriali sono in grado di reagire a stimoli ambientali specifici come la pressione, gli odori, la luce o il calore, e questo grazie ad alcuni speciali adattamenti della loro membrana. I dendriti presenti nei neuroni del cervello e del midollo spinale, reagiscono ai neurotrasmettitori chimici rilasciati da altri neuroni. La loro membrana è dotata di recettori proteici che si legano a neurotrasmettitori specifici e trasmettono, come risultato di questa unione, segnali elettrici88.
Il soma (corpo cellulare): integra i segnali elettrici provenienti dai dendriti e assicura le funzioni vitali del neurone. I segnali confluiscono al corpo cellulare del neurone viaggiando lungo i dendriti .Questo, si comporta come un centro di integrazione, "interpreta" i segnali e "decide" se produrre un potenziale d'azione: il segnale elettrico di uscita (output) del neurone. È provvisto di un'ampia varietà di organuli, come qualsiasi altra cellula organica, quindi è in grado di sintetizzare le proteine, i lipidi e i carboidrati. Funge anche da coordinatore per le attività metaboliche della cellula89.
L'assone: è la via che porta a destinazione i segnali elettrici generati dal corpo cellulare. Normalmente, l'assone, è una fibra lunga e sottile, che si protende dal corpo cellulare. Per questo motivo il neurone è la cellula più lunga del corpo umano. Singoli assoni, ad esempio, partono dal midollo spinale e arrivano alle dita dei piedi, misurando più di un metro. Gli assoni sono le linee di distribuzione, attraverso le quali si propagano i potenziali d'azione in direzione centrifuga verso le estremità del neurone. Gli assoni sono per lo più avvolti in un fascio di nervi, come i fili di un cavo elettrico. A differenza di questo, che dissipa energia nel tragitto tra la centrale e l'utente, la membrana plasmatica degli assoni fa pervenire, alle estremità del neurone, potenziali d'azione di intensità immutata, grazie, per buona parte, alla guaina mielinica che offre un ottimo isolamento90. Il segnale trasmesso è codificato in via principale attraverso la frequenza di trasmissione91.
I terminali sinaptici: essi comunicano con altri neuroni, muscoli e ghiandole. Sono come dei rigonfiamenti delle estremità ramificate degli assoni. La trasmissione dei segnali alle altre cellule avviene a livello dei terminali sinaptici. La maggior parte di essi contiene una sostanza chimica specifica, detta neurotrasmettitore, che viene rilasciata come reazione a un potenziale d'azione che percorre l'assone. I terminali sinaptici di un neurone possono essere connessi ad una ghiandola, ad un muscolo con dendriti o ad un corpo cellulare di un secondo neurone, in questo modo il segnale in uscita (output) della prima cellula può diventare segnale in entrata (input) per la seconda92.
La sinapsi: è il piccolo spazio che esiste fra i terminali di un neurone e i dendriti di un altro. Il neurotrasmettitore che viene rilasciato nella sinapsi, stimola i dendriti del neurone più vicino posizionandosi in zone specifiche della sua membrana.
Vedremo in seguito, parlando del cervello, come i neuroni abbiano un'enorme importanza per la codifica e il processo biochimico dei segnali, per ora, sapere come sono formati è sufficiente per procedere nella comprensione delle varie parti della retina.
La prima fila di cellule, nella parte posteriore della retina, è costituita dai fotorecettori: i bastoncelli e ì coni, così denominati in base all'aspetto/forma che presentano se osservati al microscopio (la prima volta vennero descritti, in maniera piuttosto approssimativa, nel 1835 da G. R. Treviranus). I bastoncelli, sono circa 120 milioni, molto più numerosi dei coniche sono circa 8 milioni. Sono responsabili della visione in condizione di ridotta illuminazione, in specie notturna, e sono fuori combattimento in piena luce. I coni non rispondono alle basse intensità luminose, ma sono responsabili della capacità di distinguere i dettagli fini, e della visione dei colori. La visione alla luce diurna, di pertinenza dei coni, viene denominata fotopica. Mentre la visione scotopica, fornita dai bastoncelli, è relativa alla luce crepuscolare. La visione mesopica, è riferita al caso intermedio, dato dalla luce lunare, che si colloca tra la brillantezza della luce solare e la debole luminosità delle stelle. Essa può risultare ingannevole93.
Se si considerano le dimensioni dei recettori retinici ci si rende condo di quanto possano essere sensibili: il più piccolo di essi misura 1 pm (picometro94), circa il doppio della lunghezza d'onda della luce rossa, dimensioni inferiori sono difficilmente ottenibili. La sensibilità, definita come la più piccola separazione rilevabile tra due linee adiacenti, date queste dimensioni, risulta molto inferiore del diametro della singola cellula a cono. Il cervello ricostruisce i gradienti di intensità in base alla quantità di segnali che riceve da un certo numero di recettori, quindi raggiunge una maggiore acuità visiva nella rilevazione delle linee che non dei punti95.
Le quantità relative di coni e bastoncelli variano in modo evidente nelle diverse zone della retina. Al centro, dove la visione del dettaglio è migliore, vi sono solo coni. Quest'area priva di bastoncelli, detta fovea, ha un diametro di circa mezzo millimetro. I coni sono presenti su tutta la retina, ma, nella fovea, sono in maggior numero e densamente stipati 96.
Soltanto i coni provvedono alla visione cromatica. Tale conclusione è frutto di numerose ricerche effettuate sugli occhi di diversi animali, analizzando la loro struttura retinica e confrontando la capacità di distinguere i colori (rilevata attraverso una serie di osservazioni sul comportamento). Infatti si è constatando che non vi sono molti coni nella periferia della retina umana, da dove non si ottiene la visione a colori. Si è osservato inoltre che, la fovea, fornisce prestazioni migliori , a livello di dettagli visivi e cromatici, ma essa, risulta meno sensibile di altre regioni della retina ricche di bastoncelli (perciò gli astronomi "escludono" la fovea quando desiderano osservare stelle molto deboli. Essi cercano di far cadere l'immagine in una regione retinica ricca di bastoncelli più sensibili. Questo, però non consente di rilevare i colori delle stelle). Queste zone ricche di bastoncelli, secondo Gregory si sono formate in stadi evolutivi remoti, per questo egli le definisce “primitive”97.
Una domanda interessante è come i fotorecettori, coni e bastoncelli, siano in grado di cambiare l'energia elettromagnetica dei protoni in attività neurale. È un processo complesso e veramente ingegnoso conosciuto in modo abbastanza completo. Sia coni che bastoncelli sono formati da due parti: il segmento interno, che contiene il nucleo e l'apparato produttivo cellulare, e il segmento esterno che contiene bilioni di pigmenti molecolari fotosensibili. Questi pigmenti molecolari sono incastonati nelle membrane di migliaia di dischi impilati come “frittelle” perpendicolari al lungo asse del segmento esterno. Il pigmento nei bastoncelli è detto rodopsina98, e si conosce come questa molecola fotosensibile converta la luce in energia elettrochimica. Quando un fotone colpisce una molecola di rodopsina e ne è assorbito, questa cambia la sua forma in modo tale da alterare il flusso della corrente elettrica all'interno e vicino al pigmento molecolare. La parte interessante è che questa complessa reazione biochimica produce dei cambiamenti nella membrana esterna del recettore. Questi cambiamenti sono poi veicolati lungo la membrana esterna fino alla regione sinaptica del recettore, dove trasmettitori chimici influenzano il neurone successivo. Le variazioni elettriche che risultano da ogni fotone assorbito dallo stesso recettore sono unite nella risposta della sua membrana esterna. La variazione complessiva del potenziale elettrico dalla parte interna a quella esterna della cellula è graduale, continuo, non come in molte altre parti del sistema nervoso. In termini quantitativi, la risposta graduale di un fotorecettore è una funzione logaritmica del numero dei fotoni assorbiti. Questo significa che la stessa crescita complessiva in uscita richiede un'aggiunta di pochi fotoni a bassi livelli ma in quantità maggiori a livelli superiori. Questa complessa catena di reazioni nel segmento esterno è denominata sbiancamento del pigmento poiché le variazioni nella forma molecolare acquisite attraverso la luce ne causano anche la variazione di colore. Prima che una molecola di rodopsina sia sbiancata dalla luce appare violacea, dopo è quasi trasparente. Poiché è trasparente, è difficile che ne assorba un altro fotone. Per riacquistare il suo stato iniziale, la rodopsina, deve attendere l'intervento di un enzima contenuto nell'epitelio dei pigmenti dietro la retina99. Diversamente dai bastoncelli, i coni hanno tre differenti tipi di foto-pigmenti, ognuno dei quali risponde a una diversa lunghezza d'onda. Sono difficili da estrarre e vengono studiati in loco con la riflessione e il densimetro. Questi tre tipi differenti di coni, con diverse caratteristiche di banda passante, sono responsabili di tre diversi canali informativi che caratterizzano la visione a colori, come accennato. Ci si è chiesti se i coni interagiscano fra loro. Nella retina della tartaruga questo è evidente. Nel caso dei vertebrati sembra che questa interattività, che porta ad associazione elettrica, tenda a ridurre il disturbo dei fotorecettori a bassi livelli di luce. È anche interessante notare che l'acuità nella visione spaziale non ne è degenerata: anzi, la comunicazione fra fotorecettori può migliorare la percezione dell'immagine ad alti livelli di luce100.
In una posizione particolare della retina, detta “macula lutea”, ci sono alcuni processi spettrali addizionali. Questa è un pigmento giallo schermato, che si trova solo nei primati, che funge da filtro per la parte blu-violetta dello spettro della luce. Questo effetto di filtraggio, aggiunto a quello effettuato dalle lenti del sistema ottico sulle onde corte, e all'assenza nella regione centrale della fovea di coni sensibili al blu, attenua, in modo significativo, la capacità di vedere le alte frequenze (ossia: radiazioni ultraviolette, raggi x e gamma)101.
Dato che coni e bastoncelli sono nella parte posteriore della retina, la luce che arriva deve attraversare gli altri due strati per stimolarli. Non è ben compreso perché la retina si sviluppi in questo modo, a rovescio. Alcuni studiosi si rifanno allo sviluppo embriologico della retina (nei vertebrati avviene a partire dalla corteccia cerebrale)102. Un'altra ragione possibile, di tipo funzionale, è, l'esistenza, dietro i fotorecettori, di una fila di cellule che contengono un pigmento nero, la melanina (che si trova anche nella pelle). La melanina assorbe la luce che ha attraversato la retina, impedendo che sia riflessa all'indietro e si diffonda all'interno dell'occhio, si comporta come la vernice nera all'interno di una macchina fotografica. Questo tipo di cellule sono anche di supporto per la ri-sintesi chimica del pigmento visivo dei recettori, precedentemente sbiancato dalla luce. È necessario, per svolgere queste due funzioni, che il pigmento melanico sia posizionato vicino ai recettori. Quindi se i recettori si trovassero nella parte anteriore della retina, le cellule pigmentate si dovrebbero collocare tra essi e lo strato successivo di cellule, in una regione dove già vi sono assoni, dendriti e sinapsi. Così, invece, gli strati anteriori ai recettori sono trasparenti e, probabilmente, non offuscano molto l'immagine. Tuttavia nel millimetro centrale, la già menzionata fovea dove maggiore è l'acuità visiva, le conseguenze di un eventuale lieve offuscamento sarebbero veramente disastrose. Questa soluzione dev'essere il risultato ottimale del processo evolutivo, che, spostando lateralmente gli altri strati ha formato un anello più spesso, ed esposto i coni centrali direttamente alla luce. La fovea risulta quindi infossata, depressa103.
Interessante è anche osservare la disposizione dei recettori retinici, che risulta a forma di matrice esagonale di coni con la presenza, negli spazi intermedi, di bastoncelli dalle dimensioni inferiori. Questo tipo di suddivisione e posizionamento, chiamato tassellamento, è ottimale, nel senso che ogni elemento ha un numero massimo di vicini equidistanti. Tale schema risulta impraticabile in un computer a scopi generici, nei quali il piano dell'immagine è suddiviso in rettangoli. Perciò, l'elemento base dell'immagine digitale nei computer, il pixel, risulta di forma quadrata104.
Il secondo strato nel nostro ordine, risalendo verso la parte anteriore dell'occhio, è posto tra coni-bastoncelli e le cellule gangliari, si tratta di una posizione intermedia della retina. Esso contiene tre tipi di cellule nervose: le cellule bipolari, le cellule orizzontali e le cellule amacrine. Le cellule bipolari ricevono l'input dai recettori, come dimostra il disegno della Figura 10. Molte di esse si connettono direttamente con le cellule gangliari. Le cellule orizzontali connettono i recettori e le cellule bipolari. Sono connessioni (spesso del tipo di molti a uno) relativamente lunghe che scorrono parallele agli strati della retina; in modo simile, le cellule amacrine connettono le cellule bipolari e alle cellule gangliari (ancora connessioni di tipo molti a uno)105.
Sulla superficie anteriore della retina c'è il terzo strato composto dalle cellule gangliari. Gli assoni di queste attraversano la retina, si raccolgono in un fascio a livello del disco ottico, quindi lasciano l'occhio andando a costituire il nervo ottico. I punti in cui i nervi ottici lasciano l'occhio sono denominati scotomi e sono zone cieche. Queste zone non costituiscono un grave problema in quanto vi è una cooperazione tra la visione bioculare e il cervello per costruire la parte mancante106. Ciascun occhio contiene circa 128 milioni di fotorecettori, suddivisi in coni e bastoncelli, ma solo un milione di cellule gangliari, quindi lo spessore del nervo ottico risultante è inferiore. Le ridotte dimensioni del nervo ottico ne aumentano la flessibilità, che, a sua volta permette un miglioramento dei movimenti degli occhi. L'enorme differenza tra i fotorecettori e le cellule gangliari pone il problema di come venga mantenuto il dettaglio nell'informazione visiva. La risoluzione sta nell'analisi delle connessioni all'interno della retina. L'informazione che attraversa la retina può seguire due strade107. La prima è una via diretta dai fotorecettori alle cellule bipolari e gangliari108. La seconda è indiretta: possono essere interposte, tra recettori e cellule bipolari delle cellule orizzontali, oppure, delle cellule amacrine tra cellule bipolari e cellule gangliari109. Il primo a descrivere queste connessioni dettagliatamente fu Ramon y Cajal intorno al 1900110. Hubel precisa che “La via diretta è altamente specifica, o privata, nel senso che uno solo, o pochi, recettori si connettono con una cellula bipolare, e solo una, o poche, cellule bipolari si connettono con una cellula gangliare. La via indiretta è più diffusa, o estesa, possedendo più ampie connessioni laterali. L'area totale dello strato posteriore occupata dai recettori che si connettono con una sola cellula gangliare dello strato anteriore, direttamente e indirettamente, è solo di circa un millimetro. Quest'area costituisce il campo recettivo111 della cellula gangliare, ossia la regione di retina la cui stimolazione luminosa modifica l'attività della cellula gangliare.”112.
Questa struttura è presente in generale in tutta la retina. Tuttavia nel dettaglio le connessioni si diversificano molto passando dalla fovea (dove è massima la nostra capacità di discriminare i fini dettagli) alla regione più esterna, la periferia113 (dove la visione diventa relativamente grossolana): cambiano le caratteristiche del collegamento tra il recettore e la cellula gangliare. Nella fovea e nelle sue vicinanze normalmente un singolo cono è connesso a una singola cellula bipolare, e, quest'ultima a una cellula gangliare. Allontanandosi dal centro, numerosi recettori confluiscono sulle cellule bipolari e queste convergono sulle cellule gangliari. Questo sistema di congiunzioni presente in gran parte della retina, assieme alla via privata della fovea e dintorni, fornisce una spiegazione di come la nostra visione risulti dettagliata anche in presenza di un rapporto di 128 a 1 tra i recettori e le fibre del nervo ottico114.
In sintesi si può affermare che parte del processo percettivo avviene direttamente nell'occhio, che risulta quindi parte integrante del cervello, poiché i fotorecettori sono interconnessi e danno luogo a "campi recettivi". Questi risultano più ampi nel caso di luce debole, in quanto è sacrificata l'acuità necessaria a distinguere i dettagli minuti per favorire una maggiore sensibilità. Questo procedimento, gestito nell'occhio a livello neurale, è assimilabile alla possibilità di optare per l'utilizzo nella macchina fotografica di una pellicola più veloce e più granulosa per le riprese effettuate con scarsa luminosità115.
Si aggiunga la presenza dell'inibizione laterale, la si potrebbe assimilare a un meccanismo neurale per rendere più definite le immagini. Funziona come segue: i fotorecettori più interessati dallo stimolo luminoso inibiscono quelli vicini meno coinvolti. Sembra, che questo meccanismo venga impiegato per filtrare il numero di segnali inviati al cervello, in quanto evidenzia i contorni, che risultano, spesso, molto significativi116.
In generale, è possibile affermare che l'occhio umano ha carattere polifunzionale e, quindi gli mancano molte delle caratteristiche specifiche di altri tipi di occhio. Ne risulta che l'informazione visiva è poco selettiva e non vincolata a speciali esigenze. Il compito di operare le scelte opportune entro un ampio ventaglio di dati relativamente neutri viene lasciato, di volta in volta, al cervello117.
Pur non essendo entrati nello specifico, se non per quanto servirà in seguito, e tralasciando alcuni aspetti di cui si parlerà più avanti, come la percezione delle distanze, dovrebbe essere fin d’ora chiaro che il processo visivo negli organismi viventi è costituito da una serie di analisi in ordine gerarchico lungo il percorso che va dagli occhi al cervello.
Se l'occhio è l'organo che si occupa di captare i segnali luminosi e le immagini, è del cervello il compito di elaborarli e, poi utilizzarli.
Passando dagli occhi al cervello, la conoscenza scientifica di strutture (anatomiche, neurobiologiche, ecc) e funzioni scema visibilmente. Questo si verifica in parte per la mancanza di adeguati strumenti d'osservazione118, e in parte si deve alla materia osservata. Per esaminare un minerale lo si può scomporre, frantumare, ecc senza alterarne la natura, ma ciò non è possibile in biologia.
Per sezionare un organo appartenente a un essere vivente bisogna innanzitutto reperirlo e, quindi, causarne la “morte”. Questo comporta problemi di natura umana, etica e sociale119. Si aggiunga che lo studio di un tessuto vivo non è paragonabile allo studio di uno morto, tra l’altro soggetto a degenerazione.
Per questi motivi, si ricorre agli animali che presentano organi simili a quelli umani, come gatti e scimmie, in specie se vi è necessità di sezionarli120. Purtroppo non si possono avere da questi delle risposte verbali. Anche se fosse possibile fare esperimenti sull'uomo vi sarebbe un grado di soggettività tale da intaccarne l’attendibilità.
Per questi motivi, la ricerca è progredita lentamente. Come per altre scienze, molto ci si aspetta dall'utilizzo del computer, in particolare dalla computer grafica, sia come strumento per visualizzare e migliorare le immagini ottenute con sonde o altri meccanismi121, sia per le sue possibilità di simulazione di eventi e comportamenti che si verificano nel corpo umano122.
Per comprendere le funzioni del cervello è necessario conoscerne, almeno in parte, la struttura fisica. Hubel afferma, probabilmente a ragione, che il modo migliore è seguire il percorso che va dall'occhio al cervello.
Dallo strato della retina di coni e bastoncelli si passa a quello intermedio di cellule neurali bipolari, orizzontali e amacrine, per arrivare a quello delle cellule gangliari i cui assoni lasciano l'occhio formando il nervo ottico, un fascio dello spessore di una matita che contiene circa un milione di fibre.
|
|
|
|
Figura 11 |
Schema del percorso visivo a opera di Gray pubblicata originariamente nel 1918. http://fr.wikipedia.org/wiki/Chiasma_optique 17 ottobre 2006 |
I due nervi ottici, come si può vedere da figura 11, s’incontrano prima di raggiungere il cervello nel chiasma ottico123. Qui le fibre che provengono dalla parte della retina vicina al naso si incrociano e passano dalla parte opposta del cervello, mentre le altre, provenienti dalla parte della retina vicina alle tempie, rimangono dove si trovano124.
Di conseguenza, anche le informazioni s’incrociano: quelle che provengono dalla parte sinistra del campo visivo vanno all'emisfero destro e viceversa. Dato che il campo visivo dei due emisferi si sovrappone, quando si guarda frontalmente, ognuno dei due emisferi ne possiede i relativi dati. Questi vengono tra loro elaborati e integrati in modo da permettere la percezione della profondità.
Successivamente, gran parte delle fibre si dirige al nucleo del genicolato laterale del talamo e poi alla corteccia occipitale (o corteccia visiva primaria). La parte minore, invece, va al collicolo superiore, che sembra occuparsi di informazioni primarie riguardanti la posizione spaziale degli oggetti e il controllo dei movimenti oculari125. Da qui molte si dirigono al nucleo del pulvinar del talamo per poi finire nella corteccia occipitale come le prime.
Il nucleo del pulvinar media i riflessi pupillari fornendo alcuni segnali di feedback dai vari stadi del processo visivo. Il talamo, che contiene anche il corpo del genicolato laterale, è una parte del cervello dove convergono tutti i segnali sensoriali ad eccezione dell'olfatto. Si suppone che in esso vi sia qualche interazione di basso livello che integri i dati raccolti dal corpo sull'ambiente.
Il corpo genicolato laterale del talamo è una tappa principale nel percorso visivo umano, in quanto una frazione significativa delle fibre ottiche si uniscono qui, come fanno altre provenienti da parti diverse del sistema nervoso centrale. È una struttura laminale (striata) che contiene sei distinti livelli di cellule. Non vi sono motivi per ritenere che le principali analisi visive si svolgano qui; nondimeno è evidente che queste cellule sono coinvolte nella visione a colori.
Vi sono inoltre delle altre fibre, che provengono dalla fovea e che attraversano il chiasma, che non passano né per il nucleo genicolato laterale, né per il collicolo superiore, ma si dirigono a una formazione reticolare, le cui funzioni non sono ben chiare, ricongiungendosi poi ad altre parti della corteccia.
Fu Ramon Y Cajal a pubblicare, nel 1893, il primo studio completo sull'anatomia del cervello umano. Circa 30 anni prima, Golgi ne aveva stabilito l’organizzazione, usando una tecnica che permetteva la colorazione selettiva di alcuni gruppi di cellule. Più tardi, verso la fine del diciannovesimo secolo, si comprese che la corteccia è divisa in aree differenti ognuna con proprie funzioni.
Come già detto, alla fine del percorso sensorio c'è la corteccia cerebrale. Vediamola più in dettaglio.126.
Il cervello non è una struttura uniforme. Esso è anzi diviso primariamente in tre parti: una mediana detta diencefalo (la cui struttura principale è il talamo) e due parti simmetriche e pari, gli emisferi destro e sinistro, che costituiscono il telencefalo. Essi sono connessi da una lamina di fibre nervose, chiamata corpo calloso 127.
|
|
|
|
Figura 12 |
Rappresentazione dei lobi celebrali e del percorso visivo. http://www.anisn.it/scuola/strumenti/visione/cortecciacol.htm 19 ottobre 2006 |
I due emisferi cerebrali si suddividono in sotto-parti, come si vede nelle figure 12-13, denominate lobi: frontale, occipitale, temporale e parietale. Si presume che ciascuno di questi abbia una funzione differente: il lobo frontale viene associato alla pianificazione, alla strategia e all'azione; il lobo parietale è implicato nella sensibilità, in particolare del tatto; il lobo occipitale controlla la visione; quello temporale si occupa sia delle emozioni, sia di alcuni aspetti della memoria.
|
|
|
|
Figura 13: |
Ricostruzione in 3D dei lobi cerebrali da posizione laterale e da sopra. Si distinguono in celeste il lobo frontale, in verde il lobo parietale, in viola il lobo temporale e in giallo-oro il lobo occipitale. Negli spaccati sono evidenziati il nervo e il chiasma ottico. http://www.biocfarm.unibo.it/aunsnc/telenf.html 7 novembre 2006 |
Le pareti degli emisferi sono costituite dalla sostanza grigia128, la corteccia cerebrale129, che contiene circa il 70 percento dei neuroni presenti nel sistema nervoso centrale umano, percentuale che ne attesta l'importanza. Per quanto riguarda le sue caratteristiche fisiche, si tratta di una lamina ripiegata che va dai 2 ai 4 mm di spessore, perfettamente inserita nel cranio. Se dispiegata, la corteccia cerebrale occuperebbe circa 1/7 m2. I neuroni sono circa 105 per millimetro quadrato di superficie, così che l'intera corteccia può essere considerata un network di 1010 di neuroni. Queste celle non sono posizionate a caso, ma sono organizzate in livelli, che alternano regioni densamente popolate con altre pressoché deserte. In una particolare regione si può notare un alto grado di uniformità delle celle130.
Si può spiegare questa struttura osservando che le fibre del nervo ottico si connettono alle cellule del corpo genicolato laterale. Gli assoni di queste ultime vanno poi alla corteccia visiva primaria. Questo insieme di connessioni ha un'organizzazione topografica: esiste una relazione sistematica (punto a punto) tra la mappa di una struttura e quella della successiva, in altre parole i diversi punti della retina, i punti corrispondenti del corpo genicolato laterale e quelli della corteccia possono essere congiunti da una linea continua. I processi nella corteccia, considerando il percorso seguito dalle fibre e i vari punti di elaborazione, risultano quindi localizzati in direzione laterale131.
È l' osservazione clinica che prova, da tempo e in modo significativo, l'organizzazione topografica delle vie visive. Quando si verifica una lesione anche di una piccola area della corteccia visiva primaria, si sviluppa una cecità localizzata, lo stesso effetto che si avrebbe se fosse lesa la parte corrispondente della retina132.
Queste scoperte sono una conquista relativamente recente, come dichiara Hubel: “Esiste dunque una mappa sistematica dell'universo visivo nel corpo genicolato laterale e nella corteccia. Negli anni Cinquanta non era del tutto chiaro il significato della mappa. A quel tempo non era accettata l'idea che il cervello eseguisse delle operazioni sulle informazioni che riceve, trasformandole in modo da renderle meglio utilizzabili. Si aveva la sensazione che la scena visiva fosse in qualche modo contenuta nel cervello: il problema era il significato che il cervello le attribuiva; forse non era un problema del cervello ma della mente.”133. Oggi, grazie anche allo stesso Hubel e a Torsten Wiesel, si è appreso che la corteccia visiva primaria trasforma profondamente l'informazione che riceve134.
È interessante scoprire che queste operazioni/trasformazioni/elaborazioni sono localizzate in punti distinti della corteccia. Il primo a formulare questa ipotesi fu Salomon Eberhard Henschen (1847-1930), seguito, successivamente da altri ricercatori. Essi mostrarono che la retina è connessa ad una zona specifica della corteccia cerebrale e non con l'intera corteccia. Questa zona del cervello fu detta inizialmente retina corticale, successivamente corteccia sensorio-visiva, e, di recente, corteccia visiva primaria o, con indicazione clinica abbreviata, area V1. In altre parole, esiste una zona specifica della corteccia cerebrale implicata nel peculiare processo della visione. Agli inizi del XX secolo Henschen faticò molto per sostenere la propria posizione, infatti molti credevano che le vie ottiche fossero connesse ad un'ampia area del cervello e che la visione non potesse essere ricondotta ad una sua parte specifica. La sua teoria era considerata un'assurda «mania di localizzazione» (localisation à outrance)135.
Studi successivi hanno comunque dimostrato che esistono più aree interessate alla visione che circondano l'area V1. Queste aree sono state denominate in modi diversi: nel contesto di questo elaborato si userà una terminologia semplificata, e cioé V2, V3, V4, V5 o MT (Medial Temporal cortex, corteccia temporale mediana) e così via (fig. 12). I numeri crescenti non indicano una gerarchia, le varie zone cooperano in parallelo alla percezione visiva. VI e V2 hanno il compito di selezionare e distribuire i segnali alle altre aree visive, mentre le altre zone sono specializzate nell'elaborazione e nella percezione di aspetti peculiari della scena visiva. A questa conclusione si è giunti solo negli ultimi 30 anni, perciò non è presente nelle prime teorie sulle funzioni e sul meccanismo visivo del cervello136.
I primi studi che considerarono la localizzazione, l'organizzazione topografica e le osservazioni cliniche delle lesioni all'area VI portarono alla conclusione che quanto era visto dall'occhio veniva interpretato dalle aree corticali circostanti (V2, V3, ecc, che hanno invece funzioni visive specifiche), definite “corteccia associativa”. La visione sarebbe quindi stata un processo essenzialmente passivo, distinto dalla comprensione137.
Questa teoria, ebbe comunque degli effetti positivi. Infatti la «retina corticale», o VI, divenne in breve l'area più studiata del cervello visivo, e forse di tutto il cervello: si è così scoperto che alla fovea (la zona centrale della retina), corrisponde una quantità enorme di corteccia in rapporto alla parte periferica della retina che risulta invece sottodimensionata in considerazione della sua estensione sulla retina stessa138. Quindi la «mappa retinica» nell'area corticale VI, se rapportata ad una comune lastra fotografica, risulta essere una trasposizione non conforme e affetta da deformazioni. Questa mappa evidenzia una parte specifica del campo visivo, così questa trasformazione viene denominata fattore di magnificazione corticale.
D’altro canto, tale teoria ha ritardato la comprensione della vera natura attiva del processo di visione, che scaturisce dall'interazione delle varie aree, le quali selezionano i dati disponibili, confrontano l'informazione selezionata con i ricordi immagazzinati e generano l'immagine, similmente al processo messo in atto da un artista139.
L'idea che le immagini del mondo visibile s’imprimano sulla retina e, una volta trasmesse, vengano ricevute e focalizzate da una parte del cervello visivo, l'area V1, per essere successivamente interpretate da un'altra area corticale, è oramai superata140.
Gran parte dei processi ha luogo in parallelo in sub-aree diverse, localizzate in zone topograficamente distinte. Ogni regione proietta delle fibre a numerose altre, anche se non a tutte. Le connessioni sono generalmente bidirezionali, cioè se l'area X è connessa con Y, Y a sua volta si connette con X. I due collegamenti non sono però completamente simmetrici in quanto si originano e terminano in livelli differenti della corteccia141. Si suppone, quindi, che la visione sia organizzata secondo un sistema modulare in parallelo, come visibile in figura 14. Curiosamente, Semir Zeki afferma che anche l'esperienza estetica è riconducibile allo stesso principio, confermando, in un certo senso, queste ipotesi142.
|
|
|
|---|---|
|
Figura 14 |
Interconnessioni bidirezionali fra aree della corteccia. Il diagramma riassume alcune delle connessioni fra aree visive di una scimmia143. |
I segnali che raggiungono l'area V1 possono riferirsi ad aspetti diversi come:al colore, alla luminosità, al movimento, alla forma, alla profondità, e così via. Le cellule che ricevono i segnali delle varie caratteristiche visive sono raggruppate in diversi livelli dell'area V1, ragion per cui si dice abbia una struttura laminare o striata. Tra questi livelli sono importanti, ai fini della visione dei colori, i cosiddetti blobs. Sono delle piccole isole, abbastanza numerose, che si caratterizzano per un elevato metabolismo, e in cui sono stipate le cellule che sono sensibili alle diverse lunghezze d'onda della luce. le varie ripartizioni specializzate di V1, successivamente, inviano i loro segnali ad altre aree visive, sia direttamente sia utilizzando una zona intermedia che circonda Vi, conosciuta come V2 (si veda ancora fig. 14)144.
Il compito dell'area V1 è dunque, principalmente di smistare i segnali visivi, come un ufficio postale centrale, inviandoli alle diverse aree visive della corteccia che la circonda. Questa operazione comporta, a sua volta, una diversa specializzazione di ogni gruppo di aree, che dipende dal tipo di segnali che esse ricevono. Perciò il “cervello visivo” è un insieme eterogeneo di molte zone, la più importante delle quali è V1. Zeki lo definisce “sistema di elaborazione specializzato”: un sistema globale dedicato ad aspetti specifici della scena visiva, che comprende sia le cellule specializzate di V1 che le aree visive altrettanto specializzate destinatarie, sia direttamente che indirettamente dei segnali inviati dalle prime145.
Poiché le singole cellule che costituiscono il cervello visivo presentano un'alta selettività al tipo di segnale o agli stimoli che ricevono ne consegue un'alta specializzazione funzionale dello stesso. Per inciso, una cellula può essere selettiva al colore e reagire al rosso, per esempio, ma non agli altri colori laddove altre cellule dello stesso tipo reagiranno in modo diverso ad altri colori. Queste cellule non risentono della direzione in cui lo stimolo si muove , è sufficiente che questo sia del colore adatto. In modo simile sono indifferenti alla forma, ossia reagiscono allo stimolo del colore adatto, indipendentemente dalla sua forma: una barra verticale o orizzontale, un rettangolo, un cerchio, un quadrato. Ancora, una cellula può selezionare un'altra qualità della scena visiva, ad esempio le linee con un'orientazione specifica, o i movimenti in una direzione stabilita, ecc. Nuovamente la selezione di un attributo specifico è associa all’indifferenza verso gli altri. Una cellula selettiva al movimento in una particolare direzione di un oggetto(cellula direzionalmente selettiva) non s'interessa né al suo colore né, di solito, della sua forma; praticamente le cellule più altamente selettive alla direzione, reagiscono in via ottimale a punti in movimento, ignorando le forme grandi e precise. Cellule che colgono lo stimolo di linee con un orientamento particolare saranno sensibili a questo, tralasciando il colore dello stimolo o dello sfondo su cui esso si staglia146.
La specializzazione è, probabilmente, fondamentale per il complesso meccanismo cerebrale diretto a cogliere l'essenza degli attributi. La questione riguarda ora in che modo questo obiettivo venga raggiunto nei singoli sistemi specializzati, a tal proposito sono state avanzate diverse ipotesi sui meccanismi neurologici alla base della costanza dell'oggetto o del colore.
La specializzazione funzionale costituisce quindi, una prima soluzione escogitata dal cervello per risolvere il problema di conoscere il mondo, ossia le sue componenti invarianti. La tipologia d' informazioni che il cervello non deve considerare per cogliere le caratteristiche principali di un oggetto, come il colore, differisce notevolmente dal tipo d’informazione che deve tralasciare per cogliere l'essenza di un altro attributo, come la dimensione. Nel primo caso è importante cogliere l'esatta lunghezza d'onda della luce proveniente solo da una superficie, nell'altro la distanza dall'osservatore. Probabilmente, a livello operativo, è più efficiente per il cervello valutare i diversi tipi di segnali in aree diverse, costruendole con un'anatomia e una fisiologia adatte a cogliere le caratteristiche principali di alcuni attributi distintivi. In sintesi, è probabile che esso abbia optato per un'elaborazione simultanea e parallela di diverse caratteristiche della scena visiva147.
Una delle prove che attestano la specializzazione delle funzioni presente nel cervello umano avviene con l'utilizzo di sistemi che individuano le variazioni del flusso sanguigno in determinate zone del cervello stesso. Nel momento in cui le cellule della corteccia reagiscono ad uno stimolo si verifica un incremento della loro attività, nel dettaglio aumenta la frequenza d'impulsi la loro emessa rispetto allo stato di riposo. L'accelerazione di questa attività provoca, a sua volta, un aumento dell’attività metabolica che, a cascata, richiede un maggiore apporto di sangue ossigenato. Alcune tecniche sofisticate permettono di convertire in immagini questo processo, quindi di misurare l'aumento del flusso sanguigno, specifico di un'area. Così si può ottenere, con relativa precisione, la posizione dell'area considerata nel cervello. Questo metodo permette inoltre, di osservare che, se un individuo normale vede una scena policroma (come una configurazione astratta in cui non sono riconoscibili gli oggetti, ad esempio le realizzazioni di Mondrian in fig. 15) le variazioni del flusso sanguigno, nelle regioni cerebrali, si limita all'area V1, in quanto è il punto in cui convergono tutti i segnali dalla retina, e a una zona esterna, il complesso delle aree V4. Invece, se la stessa persona osserva una composizione di quadretti neri e bianchi che si muovono in direzioni diverse, il cambiamento del flusso sanguigno si verifica ancora in V1 – in quanto è il centro di ricolta principale di tutti i segnali visivi – ma non in V4 (si consideri per completezza che esso si verifica in un'area esterna a V1 e distinta da V4, l'area V5). Ulteriori esperimenti hanno mostrano che altre caratteristiche della scena visiva, come il riconoscimento di volti noti, vengono elaborate in aree diverse148.
|
|
|
|
|
Figura 15 |
Due esempi di dipinti di Pieter Cornelis Mondriaan http://www.mondriaan.net/ 21 novembre 2006 |
|
Sono stati Livingstone e Hubel a proporre che le informazioni relative a colore, forma, profondità e movimento fossero elaborati in differenti percorsi neurali a partire dalla retina, evidenziando relazioni simili a quelle riportate in figura 14. Essi tracciarono il percorso che va dalle cellule gangliari retiniche (differenziato in due canali, uno per colore e forma e l'altro per profondità e movimento), passa per il nucleo genicolato laterale e arriva a V1 e V2. Da queste aree partono i collegamenti verso livelli più alti della corteccia, dove vengono elaborate informazioni specifiche: il movimento e la visione stereoscopica della profondità nell'area V5, il colore in V4, la forma in numerosi e diversi centri intermedi dell'area IT (Infero Temporal cortex, corteccia temporale inferiore). Da qui, il percorso per forma e colore porterebbe alla parte inferiore del sistema, denominata “what”, incaricata d'identificare gli oggetti, mentre il percorso relativo a profondità e movimento si dirige verso la parte dorsale/superiore del sistema, chiamata “where”, incaricata della localizzazione degli oggetti149.
La natura dei processi visivi nelle aree superiori della corteccia è meno chiara e conosciuta che non quella dell'area V1. Ad esempio, si è scoperto che alcune cellule dell'area IT si attivano alla vista delle mani di una scimmia, mentre altre dalla visione del volto. La natura dei processi spaziali che avvengono tra V1 e IT rimane comunque misteriosa. L'analisi del movimento che avviene in area MT fornisce dati all'area MST (Media Superior Temporal cortex, corteccia temporale mediana superiore) e a numerose altre aree parietali. Ma anche qui si conosce poco degli specifici processi che avvengono in questi punti terminali. Se la ricerca per quanto riguarda la corteccia delle scimmie ha fornito risultati vaghi e a volte controversi, la comprensione del funzionamento e della struttura delle aree corticali umane è anche meno chiara150.
|
|
|
|
Figura 16 |
Schematizzazione ipotetica del percorso visivo151. |
Se si è certi che le caratteristiche della scena visiva sono elaborate separatamente, non si ha prova certa che siano anche percepite separatamente. Si ritiene che nel cervello si abbia un qualche tipo di integrazione, che permette di riunire assieme i risultati delle operazioni eseguite dai distinti sistemi di elaborazione visiva, ricavandone un'immagine unificata del mondo che vediamo, un'immagine in cui tutti gli attributi sono percepiti simultaneamente. Sono state effettuate varie ipotesi su come ciò avvenga ; la via migliore per affrontare il problema in termini scientifici consiste probabilmente nel porsi la questione dell'esistenza di una “[...] registrazione temporale precisa dei risultati dell'attività compiuta dai diversi sistemi di elaborazione.”153.
Quindi, il quesito è: tutte le caratteristiche della scena visiva elaborate dalle differenti aree, vengono immesse in una registrazione sincronizzata? Se si considera un lasso di tempo relativamente lungo, superiore al secondo, vengono percepiti tutti gli attributi in sincronia. È uno stimolo per comprendere come si forma l'immagine visiva integrata. Se si considera che un secondo è costituito da 1000 millisecondi, si comprende che risulta un tempo molto lungo in termini neurologici; infatti è sufficiente un impulso inferiore a un millisecondo per attraversare una barriera sinaptica (punto di contatto tra cellule nervose) e di circa 35 millisecondi perché i primi segnali visivi arrivino alla corteccia, anche se ma la maggioranza arriva più tardi, dopo 70-80 millisecondi. Si può presumere che se li si esamina in un intervallo temporale molto breve, si potrebbe trovare il processo d'integrazione ipotizzato?
In alcuni esperimenti si sono misurati i tempi necessari a percepire il colore, la forma e il movimento, dimostrando che vengono percepiti separatamente: prima viene il colore poi la forma, quindi il movimento. Tra la percezione del colore e quella del movimento passano circa 60-80 millisecondi. Si può inferire che anche i sistemi percettivi siano in possesso di una specializzazione funzionale, e che nella visione vi sia una gerarchia temporale, sovrapposta ai sistemi di elaborazione parallela che sono diversamente localizzati. Ne consegue che se due attributi di un medesimo oggetto - per esempio il colore e la direzione del suo movimento- variano in un brevissimo lasso di tempo, il cervello coglie prima il cambiamento di colore e dopo quello di direzione, dato che percepisce il colore prima del movimento. In generale, sembra che il cervello non abbia la capacità, se si considerano intervalli temporali brevissimi, di collegare ciò che accade in tempo reale; esso, al massimo, riesce ad unire i risultati dei suoi sistemi di elaborazione. Questo implica il verificarsi di errori di collegamento in tempo reale, o quantomeno delle approssimazioni.
I risultati forniti da questi esperimenti danno importanti indicazioni sul funzionamento del cervello visivo. Dimostrano come differenti sistemi di elaborazione percepiscano l'elemento di propria competenza in tempi differenti. Questo suggerisce quindi che le funzioni di elaborazione e percezione siano svolte dagli stessi sistemi, ciò, a sua volta, permette d'ipotizzare vi sia una serie di sistemi paralleli per la percezione e l'elaborazione. I risultati delle operazioni compiute da sistemi di elaborazione separati sono le differenti percezioni. Si può quindi considerare l'esistenza di una rete di sistemi percettivi e di elaborazione distribuiti spazialmente. Inoltre, se, per definizione, la percezione è un evento conscio, si deduce che si percepisce ciò di cui si ha coscienza mentre, ciò di cui si è inconsapevoli non viene percepito. Il fatto che percepiamo due attributi, quali il colore e il movimento, in momenti separati, porta alla deduzione esistano anche consapevolezze separate, ognuna corrispondente all'attività di uno specifico sistema di elaborazione-percezione, e, che, queste distinte consapevolezze non siano sincronizzate tra loro. Quindi, per avere una percezione cosciente della scena, è necessario unire, non tanto le differenti attività dei diversi sistemi percettivo-elaborativi, ma le microcoscienze ottenute dai diversi sistemi percettivo-elaborativi.
Gli esperimenti menzionati indicano anche un'altra conclusione: i vari sistemi percettivo-elaborativi,, possiedono un elevato grado di autonomia pur interagendo tra loro. Questo è confermato anche dalla letteratura patologica in materia già accennata.
La specializzazione funzionale e la successiva integrazione dei processi suggerisce che il cervello non è un semplice cronista che limita la sua attività alla registrazione passiva della realtà fisica del mondo esterno, ma è un soggetto attivo che partecipa alla creazione dell'immagine visiva, con proprie regole e programmi.
Queste regole e programmi sono stati oggetto d’analisi in diverse scuole di pensiero ed hanno portato alla formulazione di varie teorie, di cui si farà cenno in seguito, ma prima è necessario concludere l'accenno fatto in precedenza sulla funzione e struttura delle cellule neurali.
La struttura generale delle cellule neurali è già stata trattata nel paragrafo relativo alla retina; ora è il momento di approfondire il loro funzionamento, da una parte per quel che riguarda la codifica e l’elaborazione dei segnali, dall'altra per la loro stessa diversificazione/tipologia . Questo al fine di completare quanto esposto a riguardo della corteccia cerebrale e di fornire delle indicazioni sulle reti neurali, un settore in via di sviluppo per quanto riguarda l'intelligenza artificiale.
L'importanza di comprendere il funzionamento dei neuroni sta nel fatto che questi sono l'unità d'analisi più appropriata per il sistema visivo, nel senso che si ritiene di poter spiegare la visione in termini di modelli di trasmissione che si notano dall'interazione di molti singoli neuroni. L'esistenza di un metodo per lo studio dei singoli neuroni consente agli scienziati di tracciare uno schema delle interconnessioni dell'intero sistema visivo, specificando quale operazione è compiuta da ogni singolo neurone154.
I segnali nervosi sono trasmessi sotto forma di sequenze di impulsi elettrici (detti potenziali d'azione) a frequenza modulata, i quali costituiscono la sola forma di comunicazione in entrata e uscita del cervello con il resto del corpo. È risaputo che la modulazione di frequenza offre un alto grado di stabilità e di rigetto del rumore rispetto alla modulazione d'ampiezza, il che spiega perché i neuroni funzionano in questo modo155.
Cerchiamo di meglio comprendere come avviene la generazione dei segnali e dei potenziali d'azione: i segnali dipendono dal movimento di ioni attraverso la membrana della cellula (in altre parole, dalla sua permeabilità ionica). Normalmente all'interno della cellula vi è un eccesso di ioni di potassio, mentre all'esterno vi è un eccesso di ioni di sodio. La cellula in qualche modo trattiene all'interno gli ioni di potassio e all'esterno quelli di sodio, in modo che la concentrazione di ciascuno ione ai due lati della membrana cellulare rimanga diversa. Nella cellula esiste inoltre una pompa sodica che continua a espellere ioni di sodio man mano che questi vi penetrano. In ogni caso, esiste una differenza di potenziale elettrico di circa 80 mV (microvolt) tra i due lati della membrana cellulare, e l'interno è sempre elettricamente negativo rispetto all'esterno156. Nello specifico, se si considera la parte interna di un assone a riposo, esso ha carica elettrica negativa rispetto alla superficie. Al comparire di uno stimolo - ad esempio quando un fotorecettore retinico è colpito dalla luce - la parte centrale della fibra diviene positiva, originando a un flusso di corrente che si diffonde come un'onda lungo il nervo. Quest’onda si muove con una velocità molto inferiore se rapportata a quella di un segnale elettrico lungo un conduttore: nelle fibre più ampie e spesse viaggia a una velocità di circa cento metri al secondo (360 km l'ora), mentre in quelle più sottili difficilmente raggiunge il metro al secondo (valori così bassi crearono stupore nei fisiologi del XIX secolo che ritenevano dovesse essere superiore alla velocità della luce). Le fibre più spesse, normalmente hanno un rivestimento di sostanze grasse - la guaina mielinica - che permette d'isolare ciascuna fibra da quelle vicine, oltre ad aumentare la velocità di conduzione dei potenziali d'azione157.
Il tempo di reazione agli stimoli - di circa un ventesimo di secondo - caratteristico della specie umana è dovuto in parte al tempo di propagazione lungo nervi periferici del segnale elettrico, e in parte ad un ritardo necessario alla commutazione delle sinapsi cerebrali, che si occupano di elaborare e dirigere i segnali sensoriali e motori. Come visto, questi ritardi permettono di ottenere informazioni sui differenti processi cerebrali (elaborazione di contorni, colore, forme, movimento, ecc)158.
I tempi di commutazione delle sinapsi, e quindi la velocità dell'azione potenziale, dipendono dalle proprietà della codifica di frequenza dello stimolo: la latenza e il periodo refrattario (o di refrattarietà). La latenza, o tempo di risposta effettivo, è definito come il tempo che trascorre dall'applicazione dello stimolo alla registrazione della variazione. Essa decresce all'aumentare dello stimolo. Il suo comportamento è simile per assoni di cellule diverse, anche se i tempi possono differire da cellula a cellula. Il periodo refrattario è il tempo minimo che passa tra due stimoli successivi che provocano due azioni potenziali consecutive. Infatti, una reazione della cellula è provocata solo da quegli stimoli che superano un valore di soglia. Questo valore dipende anche dal periodo refrattario. C'è un periodo morto, denominato periodo refrattario assoluto, prima del quale è impossibile avere una reazione. Passato questo periodo, il valore di soglia del secondo impulso decresce esponenzialmente all'aumentare del periodo refrattario, raggiungendo il punto in cui il neurone sarà nuovamente in grado di reagire/eccitarsi. Come per la latenza, questo periodo si comporta similmente per diversi assoni, variando solo d'ampiezza. Ne consegue che anche in caso di uno stimolo costante, gli impulsi che ne derivano saranno comunque controllati da questi due parametri, e avranno un proprio andamento. Non solo, si ricava che il modo migliore per ottenere una reazione è variare continuamente lo stimolo, e che questo si propaga solo in avanti, dato che la porzione che si è appena eccitata non può eccitarsi nuovamente finché non è trascorso il periodo di riposo159.
È importante sottolineare che non tutti gli stimoli provocano una reazione della cellula ma solo quelli che superano un valore di soglia. Infatti questo tipo di reazione può essere implementata/riprodotta artificialmente, sia dai programmi software, sia dai circuiti elettronici, come quelli denominati unità logica a soglia (ULS). Questa unità calcola una somma pesata dei suoi ingressi, confronta questa somma con un valore di soglia e produce un 1 se la soglia viene superata, altrimenti 0. In applicazioni in cui vi sono solo due azioni possibili, una singola ULS può a volte, essere in grado di calcolare l'azione corretta a partire da una appropriata codifica delle informazioni in entrata (vettore delle caratteristiche). Per problemi più complessi è necessaria una rete di tali elementi. È per questo che le reti neurali vengono così denominate, in quanto le ULS sono considerate modelli semplificati di neuroni biologici, che si attivano o meno a seconda dell'intensità e dell'attivazione delle loro connessioni sinaptiche160.
Dunque, questo è il processo che trasforma il potenziale elettrico161 presente nell'assone in una serie d'impulsi a frequenza modulata. Proseguendo è interessante vedere come questi si propagano e permettono l'elaborazione del segnale. L'assone si connette ai dendriti delle altre cellule attraverso i terminali sinaptici. Queste sinapsi sono delle connessioni chimiche che impiegano delle sostanze trasmettitrici per convogliare informazioni oltre il proprio limite. Gli impulsi di potenziale azione condotti lungo l'assone sono convertiti dalla sinapsi in potenziale elettrico nei dendriti, solitamente definito potenziale post-sinaptico (PostSynaptic Potential, PSP). Il PSP è direttamente proporzionale alla quantità di sostanza rilasciata e si satura in presenza di quantità elevate. Questo accade facilmente in quanto il tempo necessario affinché la sostanza attraversi lo spazio sinaptico è superiore al tempo intercorrente tra un impulso e il successivo. Quindi nuovo potenziale è sommato a quello rimanente del precedente impulso, facendo decadere il PSP (a seguito di latenza e periodo refrattario), portando ciò che si dice un basso potenziale. Se ne ricava che l'ampiezza di questa depolarizzazione162 dendritica è proporzionale alla frequenza media alla quale gli impulsi arrivano alla sinapsi, una forma di codifica a potenziale di frequenza.
Il tutto inizia da circa 10 neuroni diversi: sembra che ogni neurone debba proiettare una quantità elevata di sinapsi ai neuroni a sé connessi (per ogni neurone vi sono dalle mille alle centomila sinapsi di questo tipo). In ragione del gran numero di vie di stimolazione, una media di 104 fino a 105 PSP al secondo si propagano attraverso le sinapsi. Le giunzioni sinaptiche si trovano normalmente tra assone e dendriti, tuttavia vi possono essere anche tra assone e assone, tra dendriti e dendriti, tra assone e soma163.
Un aspetto interessante dal punto di vista computazionale è che possono esistere due tipi di congiunzioni sinaptiche: eccitatorie e inibitorie, che non sono però sinonime di incrementi positivi o negativi. In aggiunta, la quantità di depolarizzazione o iper-polarizzazione164 è pesata da ogni singola sinapsi, comportando una moltiplicazione parametrica. L'ampiezza del parametro può dipendere da determinate caratteristiche anatomiche come la grandezza della sinapsi e la distanza di questa dalla soma, ossia la lunghezza del dendrite. Ne risulta che l'adattamento/controllo è possibile anche prima del gap sinaptico, se l'assone si dirama nel punto terminale e vi sono due connessioni giustapposte. Questo tipo di sinapsi è più raro, e utilizzato per trasmettere un potenziale relativamente basso (rispetto alle frequenze degli impulsi in arrivo alla congiunzione), il quale controlla l'incremento della quantità di sostanza rilasciata dal terminale. Quindi aggiunte incrementali vengono utilizzate per modulare l'ampiezza del segnale in uscita dall'assone (potenziale pre-sinaptico) e il successivo “effetto domino”. Questo meccanismo viene definito “inibizione pre-sinapsi”, in quanto sono coinvolte solo le sinapsi dell'assone che portano una depolarizzazione165.
È curioso notare come questo meccanismo eccitatorio/inibitorio delle singole cellule sia riprodotto a livello generale del sistema nervoso. Le fibre nervose si raggruppano infatti per formare due sistemi nervosi chiamati ortosimpatico e parasimpatico. Questi agiscono sull'intero organismo con effetti opposti. Le fibre ortosimpatiche accelerano il battito cardiaco, mentre quelle parasimpatiche lo rallentano; le fibre ortosimpatiche diminuiscono la secrezione dei succhi gastrici, mentre quelle parasimpatiche la stimolano, e via dicendo166.
A questo punto è corretto chiedersi cosa accade alla moltitudine di dendriti uniti al soma. Un processo additivo, nel corpo cellulare, porta a una media pesata di tutti gli stimoli (inibitori e non) in entrata, che vengono convertiti nella frequenza in uscita. Quanto esposto è una semplificazione del processo, che non è ancora del tutto chiaro.
É bene riconsiderare come il sistema nervoso comunica con l'ambiente visivo esterno. L'intensità della luce è trasformata dalle cellule specifiche (gangliari, ecc), in basso potenziale elettrico. La domanda che sorge è: quali sono le caratteristiche di queste e delle altre cellule coinvolte nella visione?. La risposta viene dalle ricerche di Hubel e Torstin Wiesel, che registrarono l'attività delle singole cellule dell'area striata del cervello di un gatto quando gli ponevano dinanzi agli occhi semplici forme visive. Erano, in genere, barre luminose, proiettate su uno schermo posto di fronte al gatto. Hubel e Wiesel notarono che alcune cellule corticali del primo stadio di elaborazione (V1) davano segni di attività solo se la barra veniva posta in una determinata direzione; per inclinazioni diverse non registravano alcuna attività. Altre cellule tuttavia, rispondevano a inclinazioni diverse; alcune cellule cerebrali più interne erano sensibili a caratteristiche più generali, e si dimostravano attive indipendentemente da dove veniva stimolata la retina; altre ancora erano sensibili solo al movimento, e alcune di queste solo al movimento verso una specifica direzione. Questi esperimenti mostrano come nel cervello vi siano dei meccanismi specifici per la selezione di alcune caratteristiche degli oggetti. La percezione si realizza unendo queste caratteristiche167.
Per comprendere e avere un approccio più conciso è necessario riprendere il concetto di campo recettivo, già accennato trattando la retina. Il primo a darne una descrizione formale fu Edgar Douglas Adrian, seguito dalle ricerche di Hartline, Barlow e Stephen Kuffler168: si tratta, semplicemente, di un modello di cellule ricettrici del segnale luminoso che, a loro volta, suscitano la risposta di singole cellule sparse lungo il percorso visivo. Kuffler in particolare ne individuò due tipi: centro-on e centro-off. Egli sperimentò, ma non fu il solo, che illuminando la retina con un raggio di luce di una certa ampiezza si aveva una certa frequenza di risposta. Aumentando o diminuendo la superficie colpita si aveva sempre una diminuzione della risposta, per cui intuì che vi era una sorta di antagonismo tra una zona circolare interna (centro) e un anello circostante. Le cellule centro-on rispondono quando si illumina il centro del campo recettivo e la risposta diminuisce quando si illumina anche la parte circostante, che ha quindi una funzione inibitoria. Le cellule centro-off non rispondono quando si illumina il centro (che ha funzione inibitoria), ma quando si colpisce l'anello esterno169.
La tecnica della mappatura dei campi recettivi spiega la prima parte dell'esperimento di Hubel e Wiesel, ossia come si possono registrare delle variazioni/risposte (in gergo viene spesso usato “firing”, sparare) nella corteccia stimolando la retina.
Per quanto riguarda il resto dell'esperimento, si può dire che la scoperta, come spesso avviene, fu casuale: infatti, i due ricercatori si aspettavano che vi fossero delle reazioni a un singolo punto e non a determinate caratteristiche dell'immagine. Comunque, ulteriori approfondimenti rivelarono che vi erano tipi diversificati di cellule corticali con campi recettivi dalle caratteristiche differenti. Esse furono poi classificate in tre tipi: semplici, complesse e iper-complesse170.
Le cellule semplici furono così definite in quanto la loro risposta a uno stimolo complesso poteva essere prevista in base alla loro risposta a singoli punti di luce. Quindi il loro campo recettivo può essere individuato rilevando la risposta a singoli punti di luce che colpiscono vari punti della retina. Poiché alcune di queste cellule rispondo con più veemenza a un margine luminoso con un corretto orientamento e una corretta polarità (da chiaro a scuro o da scuro a chiaro), esse sono state spesso denominate rilevatrici di bordi. Altre hanno una lunga regione centrale che è sia eccitante sia inibitoria, con campi antagonisti da entrambi i lati, per questo rispondono bene sia a linee luminose che scure, e vengono denominate rilevatori di linee o barre. Hubel e Wiesel supposero che questi tipi di rilevatori di bordi e di linee potessero essere formati da una serie di unità centro-periferia nel nucleo genicolato laterale171.
Le cellule complesse sono le più comuni nella corteccia striata (circa 75% della corteccia striata). Sebbene anch'esse abbiano lunghi campi recettivi, differiscono dalle cellule semplici per alcuni aspetti:
Non linearità. Esse sono altamente non lineari, nel senso che rispondo poco o nulla a piccoli punti stazionari. Ne risulta che il preciso orientamento dei loro campi recettivi non può essere desunto con tecniche che misurino la loro risposta a singoli punti proiettati sulla retina.
Sensibili al movimento. Tendono ad essere altamente reattive a linee o bordi in movimento posizionati nel loro campo recettivo. Speso sono anche specializzate per direzione (sinistra-destra, alto-basso, in obliquo, ecc).
Insensibili alla posizione. Non riconoscono la posizione di alcuni stimoli. Lievi differenze nel posizionamento di una barra o di un reticolo, ad esempio, non influiscono molto sulla loro risposta.
Estensione spaziale. Le cellule complesse tendono ad avere dei campi recettivi mediamente più ampi delle cellule semplici.
La prima cellula corticale con cui Hubel e Wiesel “parlarono” attraverso il bordo della loro diapositiva era, infatti, una cellula complessa. Le caratteristiche appena esposte spiegano perché i tentativi fatti con luce diffusa, punti fissi, ecc, fossero andati a vuoto.
Hubel e Wiesel proposero che queste cellule fossero costituite dall'integrazione delle risposte di molte cellule semplici. Generalmente sembra questo il caso, tuttavia alcune ricevono direttamente gli stimoli dal nucleo genicolato laterale172.
Il terzo tipo di cellule, le iper-complesse, è stato così definito dai due ricercatori in quanto i loro campi ricettivi sono ancora più selettivi. La loro caratteristica più sorprendente consiste nel fatto che se si allunga una linea o un bordo oltre una certa lunghezza la loro risposta diminuisce, mentre risulta esser più vigorosa nel caso di linee o margini più corti. Per questo motivo vengono dette cellule end-stopped (di fine corsa). Le ricerche più recenti postulano che queste cellule siano cellule semplici o complesse con aggiunta questa caratteristica, piuttosto che una categoria a sé. In aggiunta si ritiene che il grado di “end-stopping” sia un continuo piuttosto che un fenomeno del tipo “o tutto o niente”173.
A questo punto sono state delineate per sommi capi le strutture anatomiche di occhio e cervello. E’ il momento di dare uno sguardo ad alcune teorie relative alla visione.
Nonostante si sia già accennato a differenti teorie è bene, prima di riportare quali siano le principali riguardanti la visione, operare una riflessione sul concetto di teoria. Quando si cerca di comprendere un fenomeno complesso, considerare semplicemente tutti i fatti rilevanti spesso non è sufficiente. È necessaria una teoria. Una teoria è un insieme integrato di dichiarazioni (definite ipotesi) riguardanti i meccanismi sottesi o i principi che non solo organizzano e spiegano i fatti conosciuti, ma che permettono anche di prevedere quelli che stanno per accadere.
Esse hanno tutte in comune il fatto di essere inadeguate in quanto per un dato dominio ci può essere solo una “corretta” teoria. Ma allora perché studiarle, analizzarle, ecc? La ragione sta nel fatto che comprenderne gli errori e tentare di migliorarle è un componente essenziale dell'attività scientifica, come affermavano Popper e Kuhn. Ovviamente le ipotesi di una teoria devono essere internamente consistenti, cioè non contraddirsi fra loro, pena il rigetto della teoria stesa. Non solo, esse devono essere anche esternamente consistenti, ossia non contraddire alcuni (idealmente tutti) fatti ormai assodati del dominio considerato. Perciò la teoria è considerata seriamente solo se risulta logicamente ed empiricamente consistente.
Tuttavia, può accadere che vi siano più teorie logicamente ed empiricamente consistenti che giustifichino un insieme di fenomeni (ad esempio, per la luce, la contrapposizione tra teoria ondulatoria e corpuscolare, confluite poi in una sola). Come si decide qual è la migliore? Gli scienziati credono nel principio del rasoio di Ockham. Questi dichiara che la teoria migliore è quella più parsimoniosa, cioè quella che può contare su risultati empirici con il minor numero di assunzioni.
Avendo così introdotto l'argomento, passiamo a considerare le teorie della visione, e come spesso succede, partiamo da una domanda: “Perché le cose appaiono come sono?” Tale quesito è stato posto da Kurt Koffka, illustre psicologo della Gestalt.
Vi sono molte possibili risposte, ma nell'arco del tempo sono emerse le seguenti tre argomentazioni:
La contrapposizione Ambiente – Organismo:
Con la risposta “Perché il mondo è fatto così” si prende la posizione di chi ritiene che l'analisi degli stimoli esterni sia il modo corretto per comprendere la percezione. In particolare si suggerisce che si dovrebbe esaminare il tipo d'informazione presente nello stimolo prossimo (nella retina) con lo stimolo distale (esterno, nell'ambiente)175.
Sulla stessa lunghezza d’onda le risposte “Poiché siamo come siamo” o “Poiché il nostro sistema visivo è fatto così com'è”. Tutte e tre le risposte, e le possibili posizioni intermedie, difendendo un approccio interno alla struttura percettiva, enfatizzando la natura dell'organismo piuttosto che la natura del mondo circostante.
La contrapposizione Empirico – Innato:
La risposta “Perché abbiamo imparato a vederle così” dichiara esplicitamente che la visione dipende dall'esperienza accumulata attraverso l'interazione con l'ambiente/mondo.
L'alternativa “Perché siamo nati per vedere in questo modo” afferma che vediamo in quanto il vedere è una nostra capacità innata, assicurataci dai nostri meccanismi neurali. L'apprendimento sarebbe avvenuto durante l'evoluzione a livello di specie. Il fatto che queste abilità abbiano bisogno di un periodo di maturazione non incide su questa visione, infatti essa ritiene che la programmazione genetica sia predominante.
Atomismo contro olismo:
“Dipende dal modo in cui appare ogni singola unità del campo visivo”. Con questa risposta gli atomisti sostengono che sia possibile ricostruire l'intero campo visivo mettendo insieme l'esperienza visiva di ogni singola regione.
“Dipende da come è organizzato l'intero campo visivo”. Con questa risposta “olisitca”, che si contrappone alla precedente, si considera insufficiente addizionare le singole unità, in quanto la loro valenza è data dall’insieme e nell’insieme. In questo modo la percezione di una parte sarà influenzata dalle altre. Questo approccio sottende che gli stimoli siano organizzati in modo che le proprietà di interi oggetti o scene prendano il posto di quelle delle varie regioni locali.
A questi approcci s’aggiunge quello comportamentale, che differisce dagli altri dal punto di vista metodologico. Esso, infatti, non risponde alla domanda così come formulata da Koffka il quale, ritenendo la visione un evento conscio, aveva un atteggiamento introspettivo. La domanda sottesa di stampo comportamentista è: “Che cosa ci permette di fare la visione?”. La risposta è quanto mai complessa e sarà l'oggetto delle parti successive della tesi, almeno per quanto riguarda la computer vision.
Altre teorie possono essere considerate nei vari approcci alla percezione visiva. Eccone una breve spiegazione.
In termini temporali, è una delle prime teorie riguardanti la percezione visiva. Lo Strutturalismo rientra nella scuola filosofica denominata “Empirismo inglese”, con riferimenti specifici negli scritti di Loke, Berkeley e Hume. Il padre fondatore è ritenuto Wilhelm Wundt, che la diffuse in Germania; successivamente la teoria arrivò negli Stati Uniti, grazie al suo discepolo Edward Titchener.
La teoria strutturalista propone la visione come risultato di un processo che parte da unità sensitive atomiche – definite primitive, ossia elementi indivisibili d'esperienza in una data modalità sensitiva – e arriva ad evocare ricordi di altre unità atomiche sensitive, che sono già state associate in memoria. Queste associazioni sono predisposte per avvenire ogni volta che un'esperienza sensoriale è sufficientemente vicina, nello spazio e nel tempo, ad un numero sufficiente di apparizioni. Nel caso della visione, le unità sensibili sono predisposte all'esperienza visiva del colore in ogni minuta regione localizzata nel campo visivo, che si presume provenga dai singoli fotorecettori della retina. Si ritiene che queste singole sensazioni siano unite in percezioni attraverso la semplice concatenazione: ossia, messe insieme come quando si crea un'immagine con la sovrapposizione di diverse diapositive.
L'esperienza visiva non richiama solo la memoria visiva, ma anche quella uditiva, tattile, olfattiva, e gustativa (ad esempio la mela rossa). La percezione è quindi programmata per avvenire a mezzo di processi associativi rapidi e inconsci, che vanno oltre la memoria acquisita attraverso le precedenti esperienze. Di conseguenza, come un osservatore diviene sempre più esperto del proprio ambiente/mondo attraverso meccanismi associativi, la sua percezione diventa sempre più ricca, più accurata e complessa.
Dai termini utilizzati è possibile notare l'analogia tra la teoria strutturalista e la chimica: la relazione tra semplici sensazioni e percezioni complesse riprende quella tra atomi e molecole. Ciò che unisce il tutto è l'associazione.
Tale descrizione mostra come al centro della teoria strutturalista vi sia un approccio empirista e atomista. Dal punto di vista metodologico i sostenitori dello Strutturalismo utilizzarono l'introspezione guidata. Questa consiste nel cercare di “guardarsi dentro” e osservare le proprie esperienze. Questa capacità non è comune, tanto che per acquisirla gli strutturalisti propongono un periodo di formazione guidato da un esperto.
Le grandi difficoltà incontrate dalla teoria sono strettamente legate al metodo utilizzato. Esso risulta troppo soggettivo, mentre il periodo di formazione più che migliorare i risultati li può influenzare. Per questi motivi la teoria strutturalista ha lasciato spazio alle teorie che si basano sull'analisi comportamentale.
Storicamente il movimento della Gestalt sorse come reazione allo Strutturalismo. Il termine, di origine tedesca, significa semplicemente “forma intera” o “configurazione”.
Questa teoria, come descritta originariamente nelle opere di Koffka, Wertheimer e Köhler, afferma che la percezione opera attraverso schemi innati di cui è possibile studiare le proprietà e le leggi di organizzazione. Il motto “L'intero è maggiore della somma delle parti” rende bene la posizione dei gestaltisti: la percezione è il risultato dell'organizzazione delle sensazioni, più che la loro associazione. L'intero possiede una propria struttura non rilevabile dalle singole parti che lo compongono.
Sono stati inoltre postulati dei principi di organizzazione percettiva per giustificare la probabilità con cui alcune percezioni si verificano più di altre. Alcuni di questi riguardano il modo in cui si raggruppano gli elementi delle figure, altri la separazione della figura dallo sfondo.
|
|
|
|
|---|---|---|
|
Figura 16 |
Esempi di figure che illustrano il principio di prossimità della Gestalt176. |
|
Per quanto riguarda il raggruppamento abbiamo177:
Il principio di prossimità:
Ad esempio, in figura 16, i puntini neri nella parte sinistra si organizzano percettivamente in righe verticali in quanto sono più vicini tra loro lungo l'asse verticale che non lungo l'orizzontale; viceversa nella parte destra i puntini neri formano righe orizzontali per la maggiore prossimità lungo una direzione orizzontale.
L'effetto cambierebbe se i pallini fossero neri e bianchi in quanto entrerebbe in gioco il principio di somiglianza che prevale su quello della prossimità.
Vi è poi il principio di continuazione secondo il quale la continuità delle linee viene preferita ai cambiamenti bruschi.
Infine il principio di chiusura, secondo cui le forme chiuse sono preferite a quelle aperte.
Con questi principi i punti, che di base hanno solo le caratteristiche di colore, grandezza e posizione, vengono organizzati in oggetti più complessi come linee, che hanno ulteriori caratteristiche quali lunghezza, orientamento e curvatura, confermando così il motto olistico178.
La Gestalt ha formulato degli altri assunti per la separazione di una figura dallo sfondo. Ad esempio, a parità di altre condizioni, una figura verrà percepita come simmetrica agli assi orizzontali e verticali, e preferibilmente vista come superficie più piccola all’interno di una superficie più grande, identificata come sfondo. Il tutto è sintetizzato dal cosiddetto principio della pregnanza, secondo cui tra le varie organizzazioni geometricamente possibili prevale quella che possiede la forma "migliore, più semplice e più stabile"179.
|
|
|
|
|---|---|---|
|
Figura 17 |
Immagini in cui non vi è prevalenza tra figura e sfondo: nella prima si contrappongono i volti al vaso, nella seconda l'anziana alla giovane. |
|
Talvolta si possono tuttavia costruire delle immagini in cui non vi è una chiara prevalenza figura-sfondo (fig. 17). In questi casi si alternano due percezioni a seconda di cosa si identifica come figura e di cosa si riconosce come sfondo180.
Per la Gestalt la percezione è cogliere le caratteristiche essenziali dell'oggetto che forniscono informazioni sulla sua struttura globale, piuttosto che la sua completezza o esattezza. Percepire non è un'operazione d'inferenza probabilistica o un'ipotesi, come asserisce il costruttivismo, in quanto si fonda su leggi predeterminate che strutturano l'informazione sensoriale181.
Per questo movimento l'empirismo non è alla base della percezione: ad esempio la capacità di organizzare gli oggetti non dipende dall'apprendimento per esperienza, ma sorge dall'interazione della struttura cerebrale con la struttura dello stimolo.
Se la contrapposizione tra empirico-innato è solo un argomento marginale, l’interesse dei gestaltisti per la relazione tra mente e cervello era così forte che postularono la teoria dell'isomorfismo psicofisiologico: le esperienze percettive (psicologiche) sono strutturalmente le stesse (isomorfiche) dei sottostanti eventi cerebrali (fisiologici).
Tuttavia l'isomorfismo psicofisico non è sufficiente per definire una teoria neurologica del funzionamento del cervello. Wolfgang Köhler approfondì l'argomento proponendo una prima spiegazione denominata fisica della Gestalt: un sistema fisico dinamico che converge verso uno stato di equilibrio di minima energia (portando la metafora delle bolle di sapone, indipendentemente dalla forma iniziale, evolvono sempre verso quella di una perfetta sfera). La seconda proposta, più specifica, fu che i meccanismi responsabili della percezione fossero dei campi elettromagnetici generati da eventi elettrici in milioni di neuroni. Questa spiegazione venne ritenuta plausibile ai tempi di Köhler, in quanto rispettava tutte le proprietà della fisica della Gestalt. In seguito, numerosi esperimenti dimostrarono che la distruzione dei campi elettrici non inibiva le capacità percettive. Questo portò ad un affievolimento del favore riconosciuto al movimento gestaltico, che comunque conosce un certo ritorno d’interesse nella veste delle teorie connetiviste dinamiche182.
Un'altra teoria classica della percezione visiva è quella elaborata da James J. Gibson dell'università di Cornell a New York.
Gibson accolse nella propria teoria l'enfasi sulle caratteristiche olistiche della percezione, tipiche del movimento della Gestalt. Egli riteneva, infatti, che la percezione degli oggetti non dovesse mai essere considerata separatamente dalla struttura dello sfondo su cui essi si proiettano. Per comprendere meglio la percezione di un oggetto, propose di analizzare la struttura dell'ambiente circostante l'oggetto stesso, che definì la “sua ecologia”183.
In effetti la teoria dell'ottica ecologica si basa più sulle informazioni percettive presenti nell'ambiente che sui meccanismi interni al cervello. Così facendo Gibson precede la moderna teoria computazionale.
Lo scopo della teoria ecologica era di specificare come le strutture del mondo (dovute alla riflessione della luce sulle superfici degli oggetti) illuminavano quello che egli definì matrice ottica ambientale (tutti i raggi che convergono in un determinato punto d'osservazione), permettendo così la percezione dell'ambiente attraverso il campionamento delle informazioni. In termini leggermente diversi, Gibson propose di individuare quali caratteristiche dello stimolo prossimo fornissero informazioni circa lo stimolo distale184.
Egli osservò che uno sfondo strutturato (una texture), come ad esempio un terreno o un prato, genera un gradiente di densità degli elementi che lo compongono (dimensione dei ciottoli di un selciato, grana di fili d'erba ecc.): gli elementi più vicini sono più distanziati di quelli più lontani. E’ questo gradiente di densità che dà luogo alla percezione di una superficie inclinata che recede dall'osservatore. Un oggetto che si stagli contro uno sfondo di questo tipo non viene percepito isolatamente, come se fosse sospeso nell'aria. Anzi, è proprio la parte di sfondo che l'oggetto nasconde che dà luogo alla percezione nella sua complessità, che dà informazioni sulle dimensioni dell'oggetto, sulla sua posizione ecc. Ad esempio, un albero verticale in un prato è visto come tale perché nasconde una parte del gradiente di densità del fondo, mentre la microstruttura dell'albero stesso (foglie, grana della corteccia dei rami ecc.) è costante185.
Si osservi che gli aspetti della scena visiva su cui Gibson concentrò la sua attenzione sono quelli che si mantengono invariati con il cambiare del punto di osservazione, come la presenza di un gradiente di densità nello sfondo, la verticalità dell'albero ecc. Per la teoria di Gibson è quindi cruciale considerare come si modifica la scena visiva durante il movimento dell'osservatore, e quali proprietà di questa scena rimangono invariate.
Le modificazioni complessive dell'intera scena visiva durante il movimento dell'osservatore consentono di ricavare, oltre agli elementi fissi/essenziali, le informazioni che definiscono la direzione del movimento dell'osservatore stesso186.
Gibson credeva quindi che nello stimolo retinico vi fossero sufficienti informazioni da permettere ad un organismo di percepire l'ambiente in modo non ambiguo e diretto, contrapponendosi anch'egli all'ipotesi dell'inferenza inconscia del costruttivismo (vedasi prossimo capitolo).
Sfortunatamente anche l'analisi di Gibson riesce difficilmente a ricostruire un mondo 3D da un'immagine 2D, anche con l'aggiunta della dimensione temporale dovuta al movimento. Il punto è che anche il tempo è una dimensione ambientale, e averla aggiunta al modello è ancora insufficiente per ricostruire un mondo che così possiede quattro dimensioni. Il problema della ricostruzione dell'ambiente presenta ancora troppe soluzioni possibili.
Il costruttivismo è un movimento che fonda le sue radici in vari rami del pensiero scientifico a noi contemporaneo: cibernetico, psicologico, operazionista187. È quindi, per natura, multidisciplinare. Esso incorpora in parte le teorie sulla visione appena esposte, e probabilmente per questo è oggigiorno l’approccio più seguito.
I principali assunti del costruttivismo sono188:
l'individuo partecipa attivamente alla costruzione della conoscenza;
in ogni soggetto esiste una struttura cognitiva di base che dà una determinata forma all'esperienza;
l'uomo viene visto come un sistema auto-organizzantesi che protegge e mantiene la propria integrità (autopoiesi).
Concetto fondamentale del costruttivismo è che la conoscenza umana, l'esperienza, l'adattamento, sono caratterizzati da una partecipazione attiva dell'individuo. Il reale non è considerato come qualcosa di oggettivo, indipendente dal soggetto che ne fa esperienza, ma è il soggetto stesso che lo crea, agendo attivamente per la sua costruzione. In sintesi: “L'ambiente, così come noi lo percepiamo, è una nostra invenzione.”189.
La possibilità di una conoscenza oggettiva è in dubbio. L'osservazione diretta dei fenomeni non è più considerata fonte privilegiata di conoscenza obiettiva. Quindi: “Tutto ciò che è detto è detto da qualcuno.”. Per inciso: non esistono fatti “nudi”, al di fuori delle teorie; ogni osservazione è ritenuta possibile solo alla luce di teorie, e nessuna conoscenza è data dall'ambiente, ma è sempre lo sviluppo di una conoscenza precedente190.
Per comprendere il fenomeno della visione i costruttivisti partono dal fatto che il nostro sistema visivo riesce regolarmente a ricostruire l'ambiente 3D da un'immagine 2D in modo accettabile. Dato che i dati informativi, come considerati da Gibson, sono insufficienti, la domanda è: “Com'è possibile?”. L'ipotesi dei costruttivisti è che vi sia una sorgente informativa ulteriore rispetto l'immagine retinica, che aggiunge informazioni a quelle reperite con il sistema visivo, così da individuare la più probabile soluzione al problema di ricostruzione dell'ambiente 3D fra le infinite logicamente possibili. In un certo senso il sistema visivo ricrea l'immagine191.
Le teorie costruttiviste sulla visione si basano sull'estrazione di informazioni ambientali dai modelli di stimolo retinico che Gibson descrisse nella sua teoria ecologica. Viene anche riconosciuta l'importanza delle proprietà emergenti di linee, bordi, angoli, e delle figure intere, come proposto dai teorici della Gestalt.
La teoria costruttivista della visione può essere ricondotta agli studi di Hermann von Helmholtz192, un brillante fisico tedesco, nonché matematico e fisiologo, contemporaneo di Wundt e del strutturalismo. Molte sue idee di base sono state pubblicate originariamente nel testo “Handbuch der Physiologischen Optik” (1856-1867) (Manuale di fisiologia ottica), divenuto un punto di riferimento fondamentale per gli studi sulla visione dei colori.
Una delle sue idee che perdurano nel tempo e che risulta centrale nella teoria costruttivista consiste nell’assunto che la visione dipenda da un processo di inferenza inconscia. Al contrario di Gibson, Helmholtz riconobbe il salto logico che esiste tra le informazioni ottiche direttamente disponibili dalla retina e la conoscenza percettiva che da esse deriva. Da qui propose, come appena accennato, che questo poteva essere superato attraverso delle “assunzioni” nascoste che venivano aggiunte alle immagini retiniche per ottenere delle “conclusioni” percettive relative all'ambiente. Queste assunzioni dovevano essere dei processi d'inferenza/deduttivi tali da trasformare gli insufficienti dati bidimensionali in una interpretazione percettiva corretta dell'ambiente 3D. Il processo deve essere inconscio in quanto, diversamente dai normali processi d'inferenza coinvolti nel pensiero e nella risoluzione di problemi, le persone non hanno consapevolezza di come o quando o perché o addirittura che stanno attuando inferenze visive.
È interessante chiedersi su quali basi si compiano queste inferenze. Helmholtz propose il principio di verosimiglianza: la visione arriva all'interpretazione che è più vicina allo stato dei fatti nel mondo esterno che, a sua volta, potrebbe aver causato lo stimolo retinico.
Il principio di verosimiglianza è spesso considerato antitetico al principio di pregnanza (spesso detto principio minimo) della Gestalt. Tuttavia non è semplice distinguere queste due teorie in quanto, ad esempio, somiglianza e semplicità sono spesso collegate tra loro.
Il processo che porta ad unire gli stimoli retinici, ma più in generale i dati sensori, con le inferenze e le deduzioni inconsce, risulta avere natura interpretativa euristica: il sistema visivo perpetua delle inferenze circa le probabili condizioni ambientali che possono aver prodotto l'immagine. Il processo è euristico in quanto le regole probabilistiche usate sono di natura empirica, quindi approssimative, e perciò spesso, ma non sempre, vere. Se false, portano ad errate conclusioni che si manifestano come illusioni ottiche.
Le assunzioni nascoste messe in atto dal sistema visivo sono molte e articolate. Un esempio può essere che alcuni bordi ad ampia gradazione come soffitti, pavimenti, muri e simili siano allineati con la gravitazione verticale o orizzontale ad essi. Normalmente quest'assunzione è esatta, e può servire per percepire correttamente gli oggetti che ci circondano. Se falsa, come in una stanza inclinata di un parco giochi, la nostra percezione è sorprendentemente storta, bieca. Questo succede perché tendiamo a ricondurre la situazione all'assunzione che in condizioni normali è corretta.
Un’interpretazione stretta o letterale delle inferenze inconsce porta a dire che il meccanismo della percezione si compie applicando in sequenza le regole della logica simbolica o risolvendo equazioni matematiche. Sembra che questi siano i tipi di processo supposti da Helmholtz a suo tempo. Oggi vi sono delle spiegazioni più plausibili provenienti dall'approccio computazionale. Ad esempio le reti connettive possono raggiungere conclusioni percettive basandosi in parte sui dati sensori entranti, in parte sulle assunzioni incorporate nel modello di interconnessioni tra le proprie unità neurali. Queste reti sono in grado d‘effettuare inferenze sulla base di assunzioni euristiche, senza usare la logica simbolica o equazioni matematiche.
Il paradigma dell'elaborazione dell'informazione è un concetto che assimila il cervello umano a un algoritmo di calcolo. Tale modello è stato applicato con successo non solo nella percezione visiva, ma anche a svariati fenomeni cognitivi quali la percezione uditiva, la memoria, il linguaggio, il giudizio, il pensiero e la risoluzione di problemi. L'approccio dell'elaborazione dell'informazione domina così questi temi, tanto che alcuni suggeriscono che vada a formare un “paradigma di Kuhn” per la cognizione.
Per “paradigma di Kuhn” s'intende un dato insieme di assunzioni di lavoro che una certa comunità scientifica condivide nell'operare ricerche per un determinato argomento. Le assunzioni di un paradigma comprendono normalmente presunzioni teoriche o meta-teoriche sul modo di concepire gli argomenti principali e le metodi d'approccio.
Gli sviluppi delle teorie visive hanno sempre seguito l'evoluzione tecnologica. In questo caso la spinta significativa è stata l'invenzione del computer elettronico.
La metafora del computer
L'invenzione del computer ha influenzato la ricerca teorica in due modi distinti, ma correlati tra loro:
Da una parte è divenuto lo strumento per testare le nuove supposizioni sui processi visivi e avere un riscontro su immagini reali. Questo ha fatto sorgere la computer vision come campo specifico di ricerca all'interno dell'informatica, o meglio scienza dei calcolatori. Il suo scopo è di programmare il calcolatore in modo che possa comprendere il mondo che lo circonda come fa l'uomo.
La seconda influenza è anche più rilevante: ha fornito l'analogia teorica tra processi mentali ed elaborazione dell'informazione. Per inciso: la mente è considerata come un programma che gira su una macchina chiamata cervello; la mente è il “software” e il cervello l’“hardware”.
Questo ha portato realmente a realizzare dei programmi basati su teorie percettive o, comunque, a descrivere queste teorie come dei processi di elaborazione.
Alcuni teorici hanno creduto che i fattori in comune ai binomi mente/cervello e programma/computer siano così estesi che un calcolatore adeguatamente programmato per vedere può realmente avere esperienze visive consce. Prospettiva denominata “AI forte” e contrapposta alla “AI debole”, in cui si ritiene che questo tipo di macchina stia solo simulando “processi” mentali, più o meno consci. Chiaramente è in corso un serio dibattito, con agguerrite e competenti fazioni.
Rimanendo nell'analogia tra mente/cervello e programma/computer, può risultare utile esaminare quanto si conosce dell'elaborazione delle informazioni su strumenti come i computer al fine di comprendere come funzioni la mente e rendere così più fruttuose le relative ricerche. A tal fine vengono utili le riflessioni di David Marr (1945-1980), che propose, nel suo libro, “Vision: A Computational Investigation into the Human Representation and Processing of Visual Information” pubblicato postumo nel 1982, tre livelli descrittivi per la comprensione di sistemi di elaborazione complessi: computazionale, algoritmico, e d'implementazione. Così facendo egli ha fornito un'analisi metateoretica (una teoria che indaga sulla natura delle teorie relative ad un argomento) al paradigma dell'elaborazione dell'informazione.
I tre livelli di Marr sono distinti e tutti necessari per comprendere il fenomeno:
Livello computazionale:
É il livello più astratto, definito come l'insieme di vincoli informativi disponibili per mappare/tracciare il percorso che parte con le informazioni in entrata e finisce con i risultati in uscita. Specifica, quindi, quali calcoli devono essere eseguiti e su quali informazioni, ma senza specificare come questi debbano essere realizzati. Un esempio, molto semplice, può essere un termostato per regolare la temperatura dell'abitazione collegato alla caldaia. Sopra o sotto determinati punti di soglia esso si spegne o accende.
Il livello algoritmico.
Esso è più specifico del precedente in quanto descrive come i calcoli sono effettuati in termini di operazioni del processo di elaborazione delle informazioni. Chiaramente vi possono essere modi diversi per ottenere lo stesso risultato, in questo senso il livello algoritmico corrisponde in modo più stretto al concetto di programma in campo informatico.
Per costruire l'algoritmo è necessario indicare una rappresentazione delle informazioni in entrata e in uscita, e specificare l'insieme di processi da applicare sulle stesse.
Il livello implementativo.
Questo livello descrive come realmente un algoritmo è incorporato in un processo fisico all'interno di un sistema. Così come un programma può essere eseguito su computer diversi per costruzione, così lo stesso algoritmo può essere incorporato in strumenti fisicamente differenti. Per “strumenti fisicamente differenti” sono da intendersi sia vari strumenti fisici come computer, servo-meccanismi (es. il termostato già citato), ecc, ma anche il cervello (con particolare riferimento alla neuroanatomia e neurofisiologia dei meccanismi di percezione, cognizione e azione)194.
Interessante è vedere come questi livelli descrittivi siano stati ripresi, con una prospettiva più psicologica, da Palmer e Kimchi che hanno cercato di individuare e analizzare le assunzioni implicite sottostanti alle teorie dell'elaborazione dell'informazione nella psicologia cognitiva. I livelli sono i seguenti:
Livello informativo.
La descrizione del funzionamento dei processi mentali può essere fatta in termini di eventi informativi, ognuno dei quali consiste di tre parti: l'informazione in entrata, l'operazione su questa e il risultato (l'informazione elaborata). La differenza con l'approccio di Marr sta nel fatto che qui ci si riferisce esplicitamente alla mente.
Decomposizione ricorsiva.
Questa assunzione implicita va a definire le teorie che possono rientrare nella macro area dell'elaborazione dell'informazione. In tale area non può infatti rientrare, ad esempio, la teoria di Gibson, che, pur descrivendo il funzionamento dei processi informativi, rifiuta esplicitamente di analizzare la loro rappresentazione ed esecuzione.
La decomposizione ricorsiva è essa stessa un processo che permette di scindere/specificare un evento informativo di livello superiore in un certo numero di eventi informativi che lo compongono stabilendo, inoltre, l'ordine temporale delle relazioni tra gli stessi. Il fatto che sia ricorsiva significa che può essere eseguita sui risultati fino ad ottenere degli elementi inscindibili, definiti primitive. Il problema verte sulla natura di queste; la proposta è di distinguere tra primitive “software” e “hardware”. Le prime sono un insieme di operazioni computazionali sufficienti ad eseguire un compito (livello logico). Le seconde sono quelle che riguardano operazioni che devono essere svolte da componenti fisici del sistema come i neuroni (livello fisico). È importante comprendere che per quanto si scomponga, non si arriva mai realmente al sistema fisico. Si rimane comunque a livello informativo. Questo è dovuto al fatto che l'informazione è per natura diversa dalla materia fisica o dall'energia, anche se necessita di queste per sussistere.
La principale differenza con il secondo livello di Marr è che quest’ultimo era inteso come una unità singola, a se stante, mentre qui è considerata come composta da molti livelli vicini tra loro e ordinati in via gerarchica.
L'efficacia di questo metodo sta nel fatto che permette di comprendere a poco a poco la complessità del sistema dell'elaborazione dell'informazione. Ogni successiva decomposizione rimuove la complessità implicitamente presente al livello precedente rendendo espliciti i suoi componenti.
Chiaramente questo funziona se il sistema ha una struttura gerarchica. Herbert Simon sostiene che questo sia vero per il sistema informativo umano, almeno per approssimazione. Egli parla di sistema “quasi scomponibile”. Per comprendere occorre distinguere tra le interazioni interne ai singoli componenti del sistema e quelle che avvengono tra gli stessi. Un sistema decomponibile è caratterizzato da interazioni tra componenti trascurabili se comparate con quelle interne agli stessi. In un sistema quasi scomponibile le interazioni sono deboli ma non trascurabili. L'opinione che il sistema umano sia quasi scomponibile è evidentemente contrastata dal movimento della Gestalt, ma anche altri sollevano dubbi.
Incorporazione fisica (implementazione).
In un sistema fisico l'informazione è rappresentata da stati diversi del sistema, mentre i processi (le operazioni) che utilizzano queste informazioni sono posti in essere da variazioni di stato.
È il caso di riflettere sulle modalità d'implementazione o incorporazione fisica. I componenti fondamentali di un sistema computazionale sono la rappresentazione e i processi.
La rappresentazione si riferisce ad uno stato/posizione del sistema visivo che sostituisce/significa una proprietà dell'ambiente, un oggetto, un evento; è quindi un modello di ciò che rappresenta. Da questo punto di vista, la rappresentazione è solo una occorrenza di un più vasto sistema di rappresentazioni che include due mondi legati e distinti: il mondo rappresentato esterno al sistema e il mondo rappresentante interno al sistema (denominato rappresentazione interna o semplicemente rappresentazione)195. La relazione tra i due mondi è data da una struttura similare. Spesso si tratta di una relazione di isomorfismo, una mappatura di un dominio nell'altro, così che le relazioni tra gli oggetti del primo siano riprodotte nel secondo. Più esplicitamente, l'isomorfismo sottintende una variazione causale fra i due mondi: una variazione nel mondo fisico (es: temperatura dell'aria) comporta una variazione nella rappresentazione (es: valori del termostato). Questo è importante sia per avere una rappresentazione sempre aggiornata, sia affinché questa sia attendibile e non accidentale.
Oltre ad esservi una relazione causa-effetto, per avere una rappresentazione è necessario vi sia un processo che la utilizzi. Pena l'inutilità, non esistenza, della stessa.
I processi sono i componenti attivi di un sistema computazionale che trasformano o operano sulle informazioni cambiando una informazione nella successiva. Senza rappresentazioni non vi sarebbe nulla su cui lavorare, senza processi nulla sarebbe compiuto.
Alcuni dei principali processi riguardano l'esplicitazione delle informazioni presenti nelle rappresentazioni. Ossia, i processi hanno bisogno di una rappresentazione del mondo, che tuttavia di per sé non fornisce le necessarie informazioni sull'ambiente. Ciò che serve, nel caso di un'immagine retinica, sono i limiti tra le varie regioni retiniche proiettati da superfici diverse, dove queste superfici sono localizzate in uno spazio 3D; come sono sistemate per formare oggetti significativi, e come questi potrebbero essere disposti. Tutte informazioni non evidenti nella risposta retinica allo stimolo ambientale e che devono essere estratte dalla mente/cervello.
Come suggerito da Helmholtz l'estrazione delle informazioni avviene tramite un processo d'inferenza sul risultato dell'associazione delle informazioni esterne con quelle interne.
Il processo d'inferenza parte da delle premesse per arrivare a delle conclusioni. Tralasciando le differenze che vi possono essere tra l'inferenza logica e quella percettiva, consideriamo quale sia l'oggetto di quest'ultima e come possa avvenire.
Vi sono due tipi d'inferenza: deduttiva e induttiva. La prima si caratterizza per portare a delle conclusioni certe se sono vere le premesse (es: se “Tutti gli uomini sono mortali” e “Gianni è un uomo” abbiamo “Gianni è mortale”). Se le premesse sono false, le conclusioni non possono essere provate.
L'inferenza induttiva è intrinsecamente incerta e probabilistica anche se le premesse sono vere. Un classico esempio di questo secondo tipo d'inferenza è la conclusione che le persone siano mortali. Normalmente non la si pensa una conclusione, in quanto la si usa come premessa a un sillogismo, ma su quali basi la si ritiene fondata? Non può essere dedotta dal fatto che molte persone sono morte, perché ci potrebbero essere delle persone viventi o che devono ancora nascere che non moriranno. La generalizzazione delle esperienze passate non assicura l'affidabile astrazione che tutte le persone siano mortali. L'induzione ha questo carattere d'incertezza, poiché si basa su fatti evidenti ma incompleti o probabilistici.
Molte inferenze del sistema visivo sono induttive, tuttavia possono essere ricondotte a deduttive adottando delle assunzioni nascoste. Alcuni ricercatori, come Rock e Gregory, suggeriscono che le assunzioni che portano ad una percezione veridica siano acquisite con l'apprendimento. Esse possono riguardare, ad esempio, l'effetto di convergenza di linee parallele all'orizzonte, alcuni vincoli ambientali come la gravità, ecc.
Normalmente più assunzioni sono coinvolte nella percezione della singola scena. Nella maggior parte dei casi esse concordano e portano a una conclusione comune; tuttavia vi sono casi in cui questo non accade. Si pone allora il problema d'individuare quelle corrette. Una soluzione è considerarle come vincoli deboli: informazioni restrittive che possono essere considerate ma anche sostituite da altre. La loro forza può variare, tuttavia anche le più conflittuali possono essere considerate per eventuali inferenze visive. Una possibile alternativa è utilizzare una logica fuzzy, introdotta da Lotfi Zadeh, piuttosto che una logica standard per modellare le inferenze. La logica fuzzy permette proposizioni che hanno diversi gradi di verità al posto dei normali valori “vero” e “falso”. Ulteriore possibilità è di trattare il problema come inferenza probabilistica, utilizzando il teorema di Baies. Il vantaggio di lavorare con questa struttura è il suo essere intrinsecamente probabile, e la possibilità che offre d'integrare in una singola struttura matematica diverse prove. Esso è inoltre compatibile con la logica fuzzy cosicché i due approcci possono essere utilizzati insieme.
Al di là della o delle procedure adottate, appare evidente la natura euristica di questi processi, che possono portare, ma non sempre, alla corretta ricostruzione dell'ambiente. Inoltre, l'aggiunta di vincoli di supporto (assunzioni nascoste) sembra essere l'unico modo in cui la percezione può compiersi. Le informazioni aggiunte non devono però essere dichiarate esplicitamente come regole logiche, in forma di proposizioni, come nei sillogismi. Possono anche essere incorporate in un modello di connessioni in una rete neurale.
L'ulteriore aspetto da considerare è se i processi seguano un orientamento del tipo “bottom-up” o “top-down”. Considerando la retina come il livello base e le successive tappe del percorso visivo come livelli superiori ci si può chiedere quale sia l'incidenza di ognuno sul risultato finale. Se consideriamo più influenti i primi stadi avremo una elaborazione “data-driven” ossia guidata dai dati. Al contrario, se sono i livelli superiori a correggere, in base alle assunzioni nascoste, le informazioni retiniche, l'elaborazione sarà del tipo “hypothesis-driven” o “expectation driven processing”. Intuitivamente si ritiene che la visione sia di tipo bottom-up, ma ciò si è rivelato vero solo per i primi stadi, e non per l'intero processo. Come affermato in precedenza, sia la struttura anatomica del cervello (che prevede collegamenti del tipo bi-direzionale), sia il fatto che la percezione del presente porti a delle aspettative future, al fine di poter agire in tempo, implicano una componente del sistema di tipo top-down. Ad esempio si è constatato che la rilevazione delle lettere è strettamente legata al fatto che queste appartengano a parole conosciute o insignificanti. Tuttavia non è ancora chiaro dove inizi l'influenza tra i due processi.
Dopo questa esposizione teorica dei vari approcci alla visione è opportuno porre, prima di analizzare più nel dettaglio l'aspetto computazionale, una riflessione sul linguaggio visivo.
Questo capitolo ha un carattere interlocutorio: da una parte è utile per richiamare e sintetizzare alcuni concetti presentati, dall'altra per porre alcune riflessioni e introdurre il capitolo centrale riguardante le tecniche utilizzate per la computer vision.
La prima riflessione riguarda la possibilità di considerare il sistema visivo come uno strumento per “intendere” un linguaggio, normalmente definito visivo e, prima ancora, se questo linguaggio esiste come tale, sia cioè un sistema di comunicazione con una propria struttura o grammatica197.
A tal fine è utile considerare il concetto sopraesposto di rappresentazione e vedere come quest’ultima sia presente fin dalla comparsa dell'uomo. Infatti i cacciatori del paleolitico198 ci hanno tramandato le loro incisioni sulle pareti delle caverne199 rappresentanti spesso gli animali da loro visti o uccisi.
Possono sorgere alcuni interrogativi: come fecero i loro compagni a comprendere che quelle linee rappresentavano un bisonte piuttosto che un altro animale? Come fece il disegnatore a spiegare, in assenza del linguaggio, a cosa si riferisse il disegno? E, più in generale, come possono delle linee richiamare alla memoria visiva dell'osservatore delle figure complesse? L'immagine percepita è molto diversa dal profilo che si può tracciare di un animale, o di un albero, ecc. In aggiunta le immagini dell'animale e quelle dei disegni attivano cellule retiniche diverse, e certamente stimoleranno zone cerebrali diverse. Questi stimoli, sebbene diversi, devono rievocare lo stesso “oggetto” dalla memoria. Lo stesso risultato si può raggiungere richiamando alla mente rumori e suoni tipici di un animale, oppure riproducendone la forma o le azioni con movimenti di mani e corpo. Oggi, con l'uso del linguaggio verbale e/o della scrittura, in forza delle loro proprietà simboliche e di astrazione, è sufficiente una parola, come «bisonte», per richiamare alla memoria l'oggetto-immagine e le relative caratteristiche distintive.
Il segno, o meglio il contorno, è quindi uno stimolo particolarmente efficace per il cervello. Pensare che le immagini degli oggetti del mondo esterno siano fisicamente ridisegnate e dipinte con le loro forme e colori nella nostra corteccia appare, oggi, assurdo. Esse, piuttosto, sono ridotte ad invarianti, a modelli visivi, e memorizzate come simboli dall'attività di alcune zone neurali.
Come accennato sono stati David Hubel e Torsten Wiesel a fornire la dimostrazione scientifica che i neuroni della corteccia visiva reagiscono, in modo preferenziale, al contorno degli oggetti del mondo esterno. Per questo lavoro fu loro dato nel 1981 il premio Nobel. I loro esperimenti mostrarono che le cellule neurali della corteccia sono reattivi solo a stimoli visivi rappresentati da linee o bordi con particolare orientamento e dimensioni. È molto probabilmente, a livello della corteccia, le immagini che incidono sulla retina, sono ridotte ai propri contorni o a segmenti di essi e, nell'ottica del costruttivismo, siano ricostruite da una rielaborazione di questa informazione. Si può supporre che siano queste informazioni a costituire la grammatica del linguaggio visivo.
Nel passaggio dalla retina alla corteccia il linguaggio dei neuroni evolve. Se nella retina i neuroni «vedono» l'immagine come un insieme di punti, nella corteccia i neuroni «vedono» i contorni dell'immagine.
Attualmente l'esempio più chiaro di come il cervello può simbolizzare l'informazione proveniente dai sensi è dato dalle risposte della corteccia visiva ai contorni degli oggetti, più che ad altre loro caratteristiche o attributi.
Non a caso, Ariana Fiorentini e Lamberto Maffei affermano: “Sembra quindi che i segni siano un linguaggio primitivo proprio del nostro sistema nervoso, una caratteristica che origina dalle proprietà fisiche del cervello: dalle sue connessioni, dalle sue intrinseche caratteristiche anatomiche e funzionali. Queste proprietà sono, in gran parte, innate, anche se possono essere modificate o perfezionate con l'esperienza. Sono proprietà comuni a tutti gli uomini, e costituiscono quindi, come le parole della lingua di un popolo, una base/patrimonio per comunicare.”200
I segni tracciati dal disegnatore paleolitico esprimevano quindi il vocabolario grafico del cervello, comune agli atri esseri umani del suo tempo. Le immagini erano state da loro acquisite e tradotte attraverso lo stesso linguaggio, basato sugli stessi simboli, ossia, essenzialmente dei contorni. La memoria riconduce questi ricordi alla percezione, essi si presentano del tutto simili a quelli percepiti con l'esperienza sensoriale e, quindi, facilmente riconoscibili.
Forse le origini del linguaggio segnico sono antiche quanto quelle del linguaggio della parola, collegabili alla capacità dell'uomo di emettere suoni e rumori. Secondo Noam Chomsky e altri linguisti, la struttura del linguaggio deriva dalla struttura del cervello umano. Chomsky sostiene che solo in parte il linguaggio viene appreso con l'esperienza, in quanto tutte le lingue hanno una "grammatica mentale" comune che è propria del cervello dell'uomo e che viene geneticamente ereditata. Chomsky porta a sostegno della sua teoria di come sia possibile capire frasi mai sentite prima o che si riferiscono ad eventi completamente estranei a chi ascolta. È sufficiente che la frase rispetti una determinata struttura grammaticale necessaria per legare tra loro nomi, aggettivi e verbi.
La comprensione del linguaggio avviene in quanto vi sono delle regole comuni a tutte le lingue per collegare le parole. Regole che Chomsky attribuisce a proprietà della struttura cerebrale.
Per il linguaggio parlato l'esistenza di questa proprietà è ipotetica; mentre per la visione sono disponibili prove sperimentali a supporto dell'esistenza di un linguaggio visivo neurale. Il fatto che i neuroni della corteccia visiva preferiscano certe forme piuttosto che altre potrebbe far supporre l'esistenza di un equivalente della grammatica mentale postulata da Chomsky per il linguaggio.
Con la teoria computazionale, si è visto come questa struttura possa essere ricondotta, attraverso i passaggi proposti prima da Marr e poi da Palmer e Kimchi, a proposizioni logiche implementabili in un computer, anche se vi sono ancora notevoli difficoltà dovute sia a limiti “hardware”, sia di know-how. Ciò che segue è quindi il tentativo di presentare quanto si conosce finora e di prevedere le possibili evoluzioni.
1AAVV, La bibbia di Gerusalemme, Bologna, Centro Editoriale Dehoniano, 2000 p. 35
2Cfr Arnaldo Bagnasco, Marzio Barbagli, Alessandro Cavalli, Corso di sociologia, Bologna, il Mulino, 1997 p.174
3Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, USA, MIT Press, 1999 p. 4
4Richard L. Gregory, Occhio e cervello – La psicologia del vedere, trad. it. Alberta Rebaglia, Milano, Raffaello Cortina, 1998 p. 1, (ed. originale Eye and Brain, 1998)
5Sembra un gioco di parole, ma mi riferisco alla differenza evidenziata da Denis Kaufman (Dziga Vertov, teorico e regista cinematografico russo) tra guardare, fermarsi superficialmente sugli aspetti effimeri, e vedere, ossia, partendo dall'osservazione degli stessi aspetti, trarre delle conclusioni non effimere, sostanziali.
Cfr. Carlo Alberto Pinelli, L'ABC del documentario, Roma, Dino Audino Editore, 2001 cap. 4
6Semir Zeki, La visione dall'interno – Arte e cervello, trad. it. Paolo Pagli e Giovanna de Vivo, Torino, Bollati Boringhieri, 2003 p. 21, (ed. originale Inner Vision. An Exploration of Art and the Brain, 1999)
7Cfr. Richard L. Gregory, Occhio e cervello – La psicologia del vedere, op. cit., p. 3
8Ibidem p. 9
9http://it.wikipedia.org/wiki/Evoluzione 23 agosto 2006
10Cfr. http://mmmgroup.altervista.org/i-darwin.html 23 agosto 2006
11Sono
interessanti, sull'argomento, le interviste fatte a Jéròme
Lejeune (1926-1994), genetista di fama internazionale, che dà
la propria opinione in merito al rapporto tra scienza e fede,
reperibili ai seguenti
indirizzi:
http://www.disf.org/ScienziatiCredenti/Lejeune.asp
23 agosto 2006
http://lgxserver.uniba.it/lei/rassegna/021006b.htm
23 agosto 2006
13Richard L. Gregory, Occhio e cervello – La psicologia del vedere, op. cit., pp. 35-36
14Cfr. Martin D. Levine, Vision in Man and Machine, USA, McGraw-Hill, 1985 p. XIV
15Ramesh Jain, Rangachar Kasturi, Brian G. Schunck, Machine Vision, U.S.A., McGraw-Hil, 1995 pp. 5-6
16Marco Pedroni, Giorgio Poletti, U.D. Fondamenti di Informatica, S. Bartolomeo in Bosco (Fe), Tecomproject, 1999 pp. 142-143
17David H. Hubel, Occhio, Cervello e Visione, trad. it. Enrica Maria Fava, Bologna, Nicola Zanichelli Editore, 1989 p. 14, (ed. originale Eye, Brain, and Vision, 1988)
18Ibidem p. 15
19Cfr Wolfgang Köhler, La psicologia della Gestalt, tra. it. Giannantonio De Toni, Milano, Giangiacomo Feltrinelli, 1984 pp. 13-30, (ed. originale Gestalt Psychology, 1947)
20Cfr. Martin D. Levine, Vision in Man and Machine, op. cit., pp. 6-9
21Emanuele Trucco, Alessandro Verri, Introductory Techniques for 3-D Computer Vision, New Jersey, Prentice Hall, 1998 p. 16
23http://www.funsci.com/fun3_it/protisti/mostra.htm 27 agosto 2006
24http://www.boscarol.com/pages/cs/000-colore.html 26 agosto 2006
25Cfr Wolfgang Köhler, La psicologia della Gestalt, Op. cit., pp. 13-30
26http://it.wikipedia.org/wiki/Ottica 28 settembre 2006
27Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p.22
28http://it.wikipedia.org/wiki/James_Clerk_Maxwell 27 agosto 2006
http://en.wikipedia.org/wiki/James_Clerk_Maxwell 27 agosto 2006
29http://it.wikipedia.org/wiki/Max_Planck 27 agosto 2006
http://en.wikipedia.org/wiki/Max_Planck 27 agosto 2006
30Diffusa era la metafora tra luce ed acqua, la prima si propaga nello spazio come le vibrazioni della seconda sulla superficie dello stagno; ma se per l'onda nello stagno è chiaro che è l'acqua a vibrare, nel caso delle onde luminose, cos'è che vibra?
http://www.geocities.com/codadilupo_2000/lucesuono.htm 27 agosto 2006
31Un oggetto che assorbe tutta la radiazione elettromagnetica incidente (e quindi non ne riflette). Nonostante il nome, il corpo nero irradia comunque, e deve il suo nome solo all'assenza di riflessione. Lo spettro (intensità della radiazione emessa ad ogni lunghezza d'onda) di un corpo nero è caratteristico, e dipende unicamente dalla sua temperatura.
http://it.wikipedia.org/wiki/Corpo_nero 27 agosto 2006
32Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 32
33Il
micron, abbreviazione del meno usato micrométro, è
un'unità di misura della lunghezza corrispondente a un
milionesimo di
metro.
http://it.wikipedia.org/wiki/Micrometro_%28unit%C3%A0_di_misura%29
28 novembre 2006
34L'angstrom
(Å), scritto anche ångström o angström, è
un'unità di lunghezza non appartenente al SI e pari a 10−10
metri, 0.1 nanometri o 100 picometri.L'angstrom prende nome dal
fisico svedese Anders Jonas Ångström, uno dei padri della
spettroscopia.
http://it.wikipedia.org/wiki/Angstrom
28 novembre 2006
35http://www.pd.astro.it/othersites/venere/ESO/g1.htm 27 agosto 2006
36O alla frequenza: infatti sono tra loro in rapporto costante, secondo la formula ν ·λ=3·108 m/s, dove ν è la frequenza, λ la lunghezza d'onda e 3·108 m/s la velocità della luce.
37Si ricordi il famoso esperimento di Newton che riuscì a scomporre e ricomporre lo spettro di un raggio di luce mediante prismi ottenendo sette colori: rosso, arancione, giallo, verde, azzurro, indaco e violetto.
38Cfr. Roberto Marangoni, Marco Geddo, Le Immagini digitali, Op. cit., p. 92
39http://it.wikipedia.org/wiki/Olaus_Roemer 27 agosto 2006
40Alcuni corpi si lasciano attraversare dalla luce: sono cioè trasparenti, come il vetro, certe materie plastiche, il cristallo, l’acqua stessa, purché pulita e non troppo profonda; altri, come il vetro smerigliato, la stoffa, la carta, non permettono di distinguere con precisione attraverso di essi gli oggetti, pur lasciando passare la luce: si tratta allora di corpi traslucidi. Molti altri corpi, infine, non fanno assolutamente passare la radiazione luminosa: un pezzo di legno o di metallo, un sasso, un blocco di carbone sono corpi opachi.
41http://it.wikipedia.org/wiki/Rifrazione
27 agosto 2006
http://it.wikipedia.org/wiki/Indice_di_rifrazione
27 agosto 2006
42Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 25
43Sorgenti luminose come: il sole, una lampadina, un fuoco, ecc
44Cfr. Roberto Marangoni, Marco Geddo, Le Immagini digitali, Op. cit., p. 122
45Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, USA, MIT Press, 1999 pp. 16-17
46Ibidem p. 6
47Cfr. Lamberto Maffei, Adriana Fiorentini, Arte e Cervello, Bologna, Zanichelli, 1995 pp. 75-76
48Nell'aria, la velocità del suono è di 331,5 m/s a 0ºC e di 343 m/s a 20ºC cfr. http://it.wikipedia.org/wiki/Velocit%C3%A0_del_suono 30 agosto 2006
49Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., pp. 6-7
50Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., pp. 35-36
51Un approfondimento delle relazione fra regni minerale, vegetale e animale, nonché di chimica organica si può trovare in: http://www.accademiaxl.it/biblioteca/virtuale/percorsi/testi/percorsi.asp?page=17 17 settembre 2006
52Mollusco marino commestibile, frequente lungo i litorali del Mediterraneo, dove vive attaccato a rocce in parte sommerse; ha una conchiglia di colore verdastro e di forma conica alquanto appiattita.
http://www.sapere.it/gr/DictionarySearchServlet?DS_action=ItalySearch&DS_resType=14&DS_userInput=patella 13 settembre 2006
53Considerato estinto in seguito ai ritrovamenti fossili risalenti al Paleozoico, il Nautilus, un mollusco cefalopode (con tentacoli inseriti nella parte anteriore della testa che servono da piedi) tetrabranchiato (possiede cioé due paia di branchie , anziché un solo paio come gli altri cefalopodi), è stato osservato per la prima volta in vita solamente nel 1829, pertanto è classificato come fossile vivente.
http://it.wikipedia.org/wiki/Nautilus_(mollusco) 17 settembre 2006
54Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 37
55È il gruppo di animali più vasto: comprende tutti gli organismi che non hanno uno scheletro interno. Ne fanno parte sia quegli esseri primitivi formati da poche decine di cellule, sia gli insetti più evoluti (come le formiche e le api). Ma la stragrande maggioranza degli invertebrati è composta da animali vermiformi, di ogni tipo e dimensione, che hanno colonizzato tutti gli ecosistemi terrestri. Cfr: http://www.sapere.it/tca/minisite/scienza/tuttozoologia/id100003.html 17 settembre 2006
56Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 42
57http://www.richardgregory.org/papers/copilia/curious-eye-copilia.pdf#search=%22copilia%22 20 settembre 2006
58Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., pp 43, 46,47
59Il riferimento è alla contrapposizione tra Karl Popper e Thomas S. Kuhn, se il primo sostiene che la confutabilità di una teoria consente la proposizione di successive ipotesi che, pur destinate ad essere falsificate, consentono un progresso verso la verità, il secondo afferma che nella storia c'è progresso non perché ci si avvicina a qualche meta assoluta (la verità), ma perché ci si allontana sempre più da stadi primitivi di ricerca. In altri termini nella scienza non c'è progresso “verso” qualcosa, ma “a partire da” qualcosa. Cfr Paolo Frignani, Paola Rizzati, Didattica della comunicazione, San Bartolomeo in Bosco (Fe), 2003 p. 293
60Si veda, come possibile approfondimento, gli appunti di neurofisiologia della visione con cenni di anatomia e embriologia realizzati da P.P. Battaglini per gli scopi divulgativi del Centro Interdipartimentale per le Neuroscienze B.R.A.I.N. Dell'Università di Trieste all'indirizzo
http://fc.units.it/ppb/visione/ 12 ottobre 2006
61Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., pp. 60-61
62Cfr David A. Forsyth, Jean Ponce, Computer Vision: A Modern Approach, Prentice Hall,2002 pp. 13-15
63Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 62
64Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 24
65Il minuto d'arco è una misura degli angoli e corrisponde a 1/60 di grado. Viene così denominato per non essere confuso con le misure del tempo. http://it.wikipedia.org/wiki/Primo_%28geometria%29 22 febbraio 2007
66David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 43
67Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 26
68Ibidem p.24
69Interessante la denominazione inglese “pinhole camera”, che tradotta letteralmente sarebbe camera con foro di spillo.
70Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 48
71Cfr. Lamberto Maffei, Adriana Fiorentini, Arte e Cervello, Op. cit., p. 3
72Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 20
73Ibidem
74Ibidem p. 21
75Ibidem p. 23
76Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 67
77Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., pp. 50-51
78Il pigmento è una sostanza colorata che assorbe una limitata fascia di lunghezza d'onda della luce incidente, riflettendo il resto. Il pigmento può essere organico o inorganico, biologico o prodotto chimicamente.
79Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 53
80Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 68
81Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., pp. 54-55
82Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 44
83Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 68
84http://www.etimo.it/?term=retina&find=Cerca 25 febbraio 2007
85Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., pp. 73-74
86Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 46
87Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 28
88http://www.benessere.com/salute/atlante/sist_nervoso.htm 12 ottobre 2006
89Ibidem
90Ibidem
91Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 29
92http://www.benessere.com/salute/atlante/sist_nervoso.htm 12 ottobre 2006
93Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 78
94Il picometro (simbolo: pm) è un'unità di misura della lunghezza nel SI (Sistema Internazionale di unità di misura), uguale a 10−12 metri. Altre unità di misura, con le relative corrispondenze, sono: 1 pm = 1000 femtometri; 100 pm = 1 ångström; 1000 pm = 1 nanometro.
95Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 80
96Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 46
97Gregory, seguendo la teoria evoluzionista darwiniana ritiene che si siano formati prima i bastoncelli e successivamente i coni.
Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 79
98Detta anche “porpora visiva”, è conosciuta da più di un secolo ed è stata studiata in modo esteso, in quanto può essere velocemente estratta e studiata in vitro.
Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 76
99Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., pp. 31-32
100Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 77
101Ibidem p. 78
102Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 76
103Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 46
104Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., pp. 75-76
105Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 47
106Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 84
107Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 47
108Ibidem
109Ibidem
110Ibidem
111In senso proprio, il termine campo recettivo si riferisce semplicemente ai recettori specifici che fanno capo a una data cellula del sistema nervoso, con l'interposizione di una o più sinapsi. Attualmente il termine tende a includere la partizione del campo o, meglio, tende a dare una indicazione di come bisogna stimolare una regione per far rispondere la cellula. Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., pp. 52-53
112Ibidem p. 48
113Si possono distinguere la fovea vera e propria di circa 0,3 mm, la “parafovea” di circa 2,5 mm, la “perifovea” con l'anello interno di circa 2,5 mm ed esterno di 5,5 mm e infine la “retina periferica” che costituisce il 97,25 percento della superficie concava della retina.
Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., pp. 73-75
114Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., pp. 47-49
115Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 78
116Ibidem
117Ibidem p. 80
118Ad esempio il già citato microscopio, pur essendo presente in forme rudimentali da tempi immemori, è stato perfezionato nel 1590, quindi relativamente di recente, da due olandesi, Zaccharias Janssen e suo figlio. Essi osservarono che aumentando il numero delle lenti in un tubo e regolandone la distanza e la grandezza, l'oggetto osservato diventava incomparabilmente più grande. Era l'antesignano del microscopio composto e del cannocchiale sviluppato poi da Galileo in Italia.
http://www3.iperbole.bologna.it/bologna/malpighi/mostra/microsco.htm 17 ottobre 2006
119In passato, per studi di anatomia, venivano usati i corpi dei defunti, con preferenza per donne e delinquenti in quanto si riteneva non avessero l'anima. Inoltre chi esercitava questo tipo di studi veniva spesso perseguitato; anche Leonardo da Vinci ebbe di questi problemi.
http://it.wikipedia.org/wiki/Anatomia_umana#Storia_dell.27anatomia 17 ottobre 2006
http://it.wikipedia.org/wiki/Leonardo_da_Vinci 17 ottobre 2006
http://www.sapere.it/tca/MainApp?srvc=dcmnt&url=/tc/scuola/percorsi/DP_anatomia/anatomia1.jsp 17ottobre 2006
120Contro la procedura di vivisezione esistono numerose associazioni e si è giunti ad alcuni progetti di legge, si veda:
http://www.novivisezione.org/ 17 ottobre 2006
121Ad esempio le recenti tecniche di scansione PET (Tomografia ad emissione di positroni), NMR C(Risonanza Magnetica Nucleare) e CT (Tomografia computerizzata). Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 98
122Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. XII
123La denominazione deriva dal fatto che assomiglia alla lettera greca x, pronunciata “chi”.
124Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 80
125Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 35
126Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 84
127http://www.sapere.it/tca/MainApp?srvc=vr&url=/5/307_1 03 novembre 2006
128Così denominata in quanto è il colore che assume nei cervelli non vivi conservati.
129Nell'uso corrente, tutta la massa nervosa contenuta nel cranio, che invece è detta più propriamente encefalo, di cui il cervello costituisce la parte anteriore e superiore. Oltre al cervello, infatti, l'encefalo comprende il tronco cerebrale (bulbo, ponte di Varolio e mesencefalo) e il cervelletto. http://www.sapere.it/tca/MainApp?srvc=vr&url=/5/307_1 03 novembre 2006
130Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 84
131Cfr David H. Hubel, Occhio, Cervello e Visione, Op. cit., p. 70
132Ibidem
133Ibidem
134Ibidem
135Cfr. Semir Zeki, La visione dall'interno – Arte e cervello, Op. cit. pp. 31
136Ibidem p.32
137Ibidem
138Ibidem p.34
139Ibidem p. 38
140Ibidem pp. 79-80
141Cfr. Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 40
142Cfr. Semir Zeki, La visione dall'interno – Arte e cervello, Op. cit. pp. 80
143Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 42
144Cfr. Semir Zeki, La visione dall'interno – Arte e cervello, Op. cit. pp. 80
145Ibidem pag. 82
146Ibidem pag. 84
147Ibidem pag. 84-85
148Ibidem pag. 87
149Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 43
150Ibidem
151Ibidem
152Cfr. Semir Zeki, La visione dall'interno – Arte e cervello, Op. cit. pp. 88-91
153Ibidem p. 89
154Stephen E. Palmer, Vision Science – photons to phenomenology, Op. cit., p. 64
155Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 85
156Cfr. Isaac Asimov, Il libro di Biologia, trad. it. Alessandra Fois, Milano, Arnoldo Mondadori, 1987 p. 351 (tit. originale Asimov's guide to Science 1984)
157Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 103-104
158Ibidem
159Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 89-90
160Cfr Nils J. Nilsson, Intelligenza artificiale, trad. it. Salvatore Gaglio, Apogeo,2002, p. 47-48 (ed. originale Artificial Intelligence: a New Synthesis, 1998)
161Il
potenziale elettrico, noto anche con il nome di potenziale scalare,
viene indicato dalla lettera V, o a volte anche dalla greca φ.
Esso si ricava a partire dal lavoro del campo
elettrico.
http://it.wikipedia.org/wiki/Potenziale_elettrico
12 dicembre 2006
162La depolarizzazione è una variazione/escursione positiva rispetto allo stato di riposo del neurone situato a circa -60 mV, quindi normalmente negativo.
163Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 91-92
164Escursione negativa, si veda la nota precedente relativa alla depolarizzazione.
165Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 91-92
166Cfr. Isaac Asimov, Il libro di Biologia, Op. cit. p. 347
167Cfr Richard L. Gregory, Occhio e cervello – La psicologia del vedere, Op. cit., p. 110
168Cfr. Martin D. Levine, Vision in Man and Machine, Op. cit., p. 102
169Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit., p. 147
170Ibidem p. 151
171Ibidem
172Ibidem p. 153
173Ibidem pp. 153-154
174Il paragrafo che segue è quasi tutto tratto dal cap. 2 di Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit. a cui si rimanda.
175Si veda il paragrafo sulla retina.
176Ariana Fiorentini, Lamberto Maffei, Arte e cervello, Zanichelli, Bologna, 1995 p. 9
177Ibidem
178Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit. p. 51
179Ariana Fiorentini, Lamberto Maffei, Arte e cervello, Zanichelli, Bologna, 1995 p. 10
180Ibidem
181Ibidem
182Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit. p. 51-53
183Ibidem
184Ibidem
185Ariana Fiorentini, Lamberto Maffei, Arte e cervello, Zanichelli, Bologna, 1995 p. 11
186Ibidem
187http://www.oikos.org/voncostrutt.htm 20 dicembre 2006 di Ernst von Glasersfeld
188http://it.wikipedia.org/wiki/Costruttivismo_%28psicologia%29 20 dicembre 2006
189Ibidem
190Ibidem
191Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit. p. 56
192http://www.anisn.it/scuola/strumenti/visione/helmholtz.htm 20 dicembre 2006
193Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit. pp. 70-85
194http://kybele.psych.cornell.edu/~edelman/marr/marr.html 23 dicembre 2006
195Stephen E. Palmer, Vision Science - Photons to Phenomenology, Op. cit. p. 77
196Cfr. Ariana Fiorentini, Lamberto Maffei, Arte e cervello, Zanichelli, Bologna, 1995 pp. 46-48
197Cfr. Giorgio Graffi, Segio Scalise, Le lingue e il linguaggio Introduzione alla linguistica, Bologna, Il Mulino, 2003 p. 16
198Letteralmente età della pietra antica che va da circa 2,5 milioni di anni (in Africa) a 10 mila anni fa. http://es.wikipedia.org/wiki/Paleol%C3%ADtico 28 dicembre 2006
199Tra le più conosciute quelle di Altamira in Spagna. http://www.artepreistorica.it/links/altamira.asp 28 dicembre 2006
200Ariana Fiorentini, Lamberto Maffei, Arte e cervello, Op. Cit. p. 48






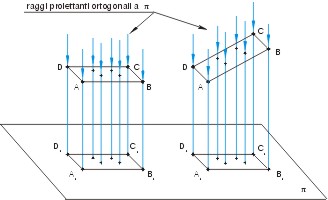

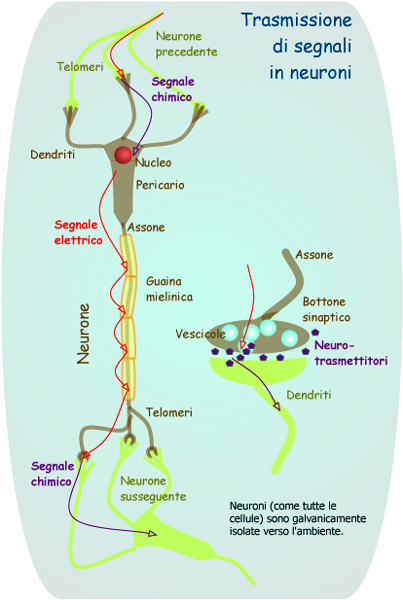












{kind=link}
{kind=link}